Introduction
Climate change is one of the main factors that may directly or indirectly influence the physiology of plants. Climate change projections suggest 0.3℃ to 4.8℃ increase of global average temperature by the end of the 21st century with a likely increase of at least 1.5℃ (IPCC, 2013). Global climate has been changing due to human activities like emissions of greenhouse gases cloudiness, aerosols, and land use changes in recent decades. Changes in atmospheric CO2 levels, temperature, rainfall, and other weather conditions affect the dynamics of weed species, distribution and their competitiveness within weed populations and with other crops across all over the world. This may help for adaptations in crop management practices which in turn will affect weed growth or the proliferation of some species.
Rice is one of the most important crops in the world. Among the major crop species, rice is the most stable food crop in Asia where more than 90% of the world rice is produced. Rice is always a vulnerable crop to climate change in terms of variation of yield and quality (Wassmann et al., 2009). Rice alone contributes a major share of the dietary requirement for the one-third population in the world. It was predicted that climate change may have a positive or negative impact on rice production irrespective of the area (Horie et al., 2000). Among the different growth stages of rice, few stages are very sensitive to temperature. Variation of temperature during growth stages highly influenced on yield and it could end up in different vegetative and reproductive outcomes (Horie et al., 2000).
Increasing temperature influences on both crops and weeds. Crop performance is closely related to the weed growth inhabiting adjacently. Weeds are the serious factor of the yield loss in rice production in the world. Rice yield loss due to the weed infestation is around 10 percent of total production throughout the world (Bastiaans and Kropff, 2003). The rice yield reduced from 40 to 100% due to the weed infestation in Korea (Kim and Ha, 2005). Both annual and perennial weed species are the most dominating pests in paddy fields in Korea during the past two decades. Presence of perennial weeds has increased from 19% in 1971 to 54% in 1981 and 60% in 1990. Park et al. (2002) reported that the most dominant weed species M. vaginalis, E. crus-galli, L. prostrata, S. planiculmis, E. kuroguwai, and S. sagittifolia have been found in China, Japan, and Korean paddy fields. Although, these weeds were originated in different locations, they are mostly visible in every rice fields of these countries.
Many studies have been undertaken to investigate the ecophysiological effects of elevated temperature and CO2 on crops (Jones and Thornton, 2003; Tubiello et al., 2002). Most of the previous studies have been conducted to investigate the effects of elevated temperature on crops, e.g. the effects of elevated temperature on rice yield and growth during the last several decades (Baker, 2004; Baker et al., 1992; Horie et al., 2000; Imai et al., 1985; Kim et al., 2003; Sakai et al., 2006; Sasaki et al., 2007; Yang et al., 2006; Yoshida, 1973a, b; Ziska et al., 1996). Most of the weed species in rice cultivation showed their dominance in machine transplanted paddy fields (Chae and Guh, 1999) than direct water-sown rice field (Kim and Pyon, 1998) and the occurrence of perennial weed species were in temperate rice fields (Kim, 1983).
However, studies on the growth response of rice and dominant weed population under elevated temperatures have not been investigated in details yet. Climate changes may vary with regions and might influence differently on rice and weed growth. The growth of weed population and rice cultivation may have diverse effect under elevated temperature regimes. So, it is necessary to conduct research regarding on rice growth due to the competitiveness with paddy weeds under elevated temperature conditions in many areas. Here, we investigated the growth response of rice crops as well as annual and perennial weed species those are dominant in Korean paddy fields under elevated temperatures.
Materials and Methods
Experimental site and manipulated temperature in the field phytotrons
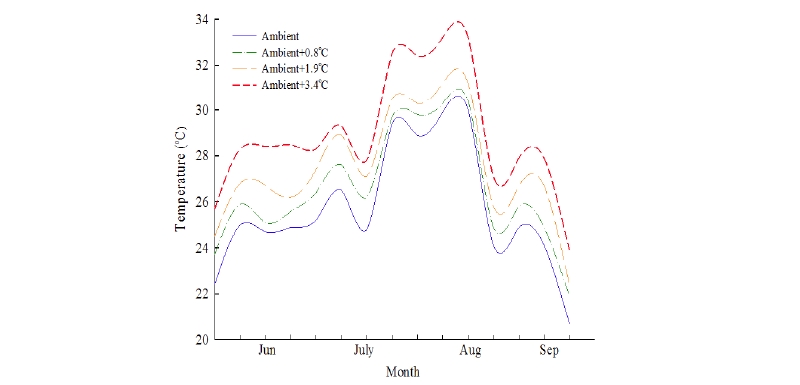
The experiment was carried out at the experimental farm in Chungnam National University, Daejeon, Korea (127°21′ E, 36°22′ N, alt. 34 m) in 2015. Rice (O. sativa), three annual (L. prostrata, M. vaginalis, and E. crus-galli) and three perennial (S. planiculmis, E. kuroguwai, and S. sagittifolia) weed species were grown in four phytotrons, each in a unit plot of 20 m × 5 m were built by establishing semi-circular (tunnel-shape) steel frame on the ground and then covered by thin clear plastic sheet to prevent rain and elevate temperature inside the phytotron. A temperature control device was engineered by sensors to automatically open and close the ventilation of plastic windows to pre-calculated variable apertures to allow for the exchange of air that maintained the desired elevation of temperatures (ambient and, 0.8, and 1.9, and 3.4 °C above ambient level). One phytotron was maintained at an ambient temperature as a control. Inside of each phytotron, a heater was installed for maintaining temperature and set up a digital temperature control box with internet mobile based control system to provide the necessary temperature for rice and different paddy weeds. The elevated temperature level was continuously monitored using an internet mobile based control system. Temperatures were measured in every 10 seconds by sensors in the phytotron and values were stored inside a smart cell phone system. Phytotrons were set to maintain temperatures at different levels i.e., at ambient, ambient+0.8°C, ambient+1.9°C, and ambient+3.4°C. The actual monthly mean temperatures in the four phytotrons during the rice and weed growing season were 25.7℃, 26.5℃, 27.4℃, and 28.9℃ which were 0℃, 0.9℃, 1.8℃, and 3.2℃, respectively above ambient temperature (Fig. 1). Phytotrons achieved their purpose as rice and weeds adapted quickly and established successfully under phytotrons-generated temperatures. The study was done by completely randomized design (CRD) with four replications.
Plant materials
Seeds of rice cultivar (Dongjin 1), three annual, and three perennial weed seeds were collected from previously harvested in the experimental field at College of Agriculture and Life Sciences, Chungnam National University, Korea. Rice and weed seeds were placed to germinate on Petri dishes with filter paper wetted with water in a controlled growth chamber maintaining the temperature of 30℃ at day and 25℃ at night for two weeks. It was considered as germinated when the radicle emerged through the seed coat. Seeds of rice cultivar and weeds were sown in plastic trays (one seed per cell) filled with upland paddy field soil. Rice and weeds seedlings were raised inside of each phytotron based on different temperatures. Four weeks after sowing, rice and weed species were transplanted to plastic pots (one plant per pot) at 2-3 leaf stage. The size of each plastic pot was 20 cm inside diameter, 17.0 cm height, 0.2 cm thickness, and filled with a mixture of commercial potting soil and sand. The total number of experimental pots was 560. In each phytotron, a total of 140 pots were used for seven plant species with four replicates which were arranged randomly with 40 cm space among each other. After that different temperature regimes were maintained as per experimental specification and the water volume was maintained up to 3±1 cm by 2 to 3 time’s top irrigation in a day.
Sampling and harvesting of plants
The plant height, dry weight, and leaf area were recorded every 10 days following sowing time to until 110 days. Rice and all weed species were collected by plucking them from ground level (except root) and transferred to the laboratory immediately after collecting for recording necessary parameters. For dry matter assessment plant samples, both rice and weeds were softly wrapped with clean paper towels and placed in an electric oven drier at 72°C for three days prior to measuring the dry weight with an electric balance. The leaf area was determined with an electronic planimeter (Li-3100C with LI-3050 conveyer accessory, LI-COR, Daejeon, Korea).
Statistical analysis
The data were tabulated as mean ± standard error for growth parameters (plant height, dry weight, and leaf area) of rice and weeds and were analyzed by using the one-way analysis of variance (ANOVA) procedure with SPSS 17.0 (SPSS, Chicago, IL, USA). The differences between the means of the treatments were adjusted by using Duncan’s multiple range test. In all the analyses, significant effects were declared at P<0.05.
Results and Discussion
It was observed that all plant species responded positively to the elevated temperature while the growth rate of weeds was higher than that of rice and the highest increase of plant height, dry weight, and leaf area were obtained at the ambient+3.4℃ (Table 1). Rice plant height was increased by 17.2% at the condition of ambient+3.4℃ than it was at the ambient temperature condition. The plant height of annual weeds i.e. E. crus-galli, M. vaginalis, and L. prostrata were 87.6%, 39.3%, and 32.9% higher, respectively at the ambient+3.4℃ than they were at the ambient. E. crus-galli showed the highest increase of plant height followed by M. vaginalis at the ambient+0.8℃, +1.9℃, and +3.4℃ compared to other weed species and rice (Table 1). In the case of perennial weed species, the plant height of S. planiculmis, E. kuroguwai, and S. sagittifolia were 26.1%, 16.9%, and 81.8% higher, respectively at the ambient+3.4℃ compared to the ambient condition. It is mentionable that S. sagittifolia showed the highest increase rates of plant heights at all elevated temperature conditions compared to other weed species and rice (Table 1). Elevated temperature created an environment to induce the growth of plant height rapidly in rice and weeds which might be occurred due to its adaptation to the environment.
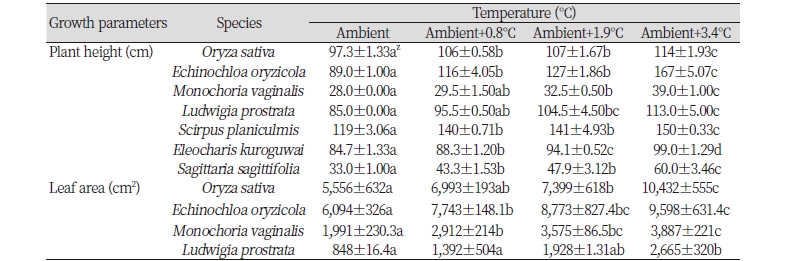
Leaf area of rice was 87.8% higher whereas E. crus-galli exhibited 57.5% higher at the ambient+3.4℃ than that of the respective control at the ambient, which was the smallest increase rate at the ambient+3.4℃ among other plants. Leaf areas of the other two weed species M. vaginalis and L. prostrata were 95.2% and 214.3% higher, respectively at the ambient+3.4℃ compared to the ambient temperature. Among the weeds species, L. prostrata showed the highest increase rates of leaf areas at all elevated temperature conditions (Table 1). Therefore, there was a strong positive relationship between growth parameters observed at different growth stages and temperature elevation. Daily rate of the increase in leaf area per plant appeared to be ruled mainly by the rate of leaf expansion of individual leaves where water and assimilates were not limited which was also largely influenced by temperature. The leaf area usually depends on the numbers and sizes of leaves, both of which are influenced by the effects of temperature on leaf initiation and rate of expansion.
A rapid increase of dry weight of rice and annual and perennial paddy weed species has been detected under elevated temperatures based on the non-linear regression analysis of accumulative dry weight of rice, three annual and three perennial weeds (Fig. 2, 3). The estimated maximum dry weight of rice at the ambient +3.4℃ was 30.4% higher while dry weight of E. oryzicola, M. vaginalis, and L. prostrata were 78.2%, 171.4%, and 211.8% higher, respectively at the ambient+3.4℃ than that of the ambient condition. L. prostrata showed the highest increase rates of dry weight at the ambient+3.4 and +1.9℃ (Fig. 2). Likewise, dry weights of S. planiculmis, E. kuroguwai, and S. sagittifolia were 38.1%, 63.2%, and 124.8% higher, respectively at the ambient+3.4℃ compared to their respective control at the ambient. S. sagittifolia showed the highest increase rates of dry weight at ambient+3.4 and +1.9℃ (Fig.3).
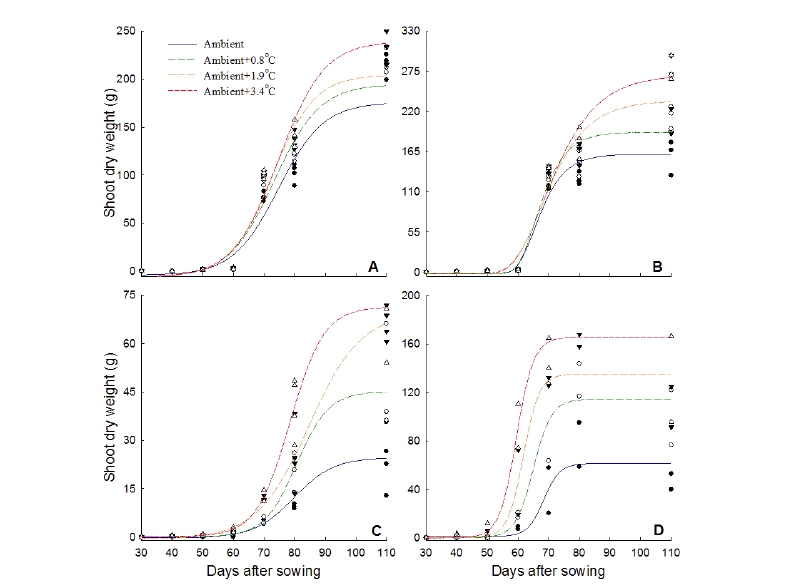
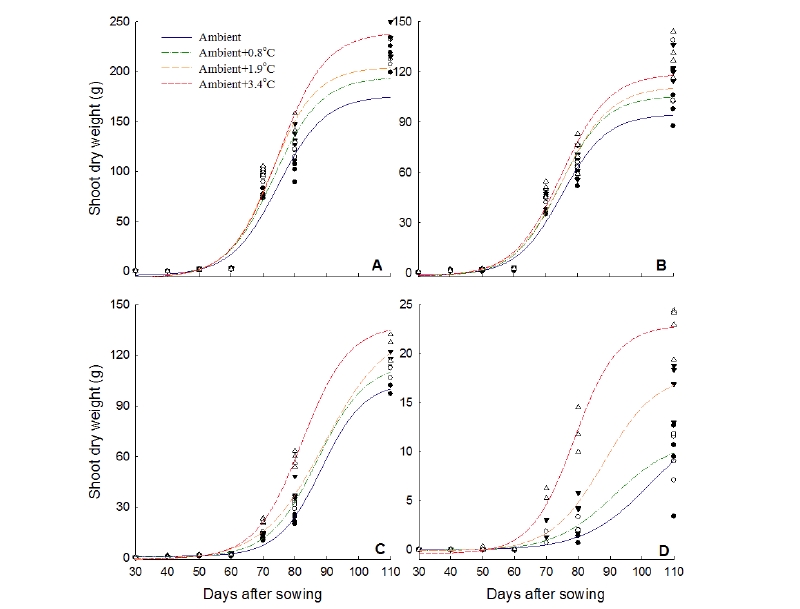
From a previous study in Korea, it was reported that seedling emergence of M. vaginalis and S. juncoides took place 1-2 days earlier and their growth took place 3 days earlier under elevated temperature (Park et al. 2010). In another study, Won et al. (2016) proclaimed that rice and three major paddy weed populations and their growth pattern response was altered under elevated temperature conditions in Korean paddy fields. Inhibition of growth with the increase of temperature often occurs through induction of ascorbic acid (ABA) as reported by Essemine et al. (2010). Furthermore, Kumar et al. (2011), as well as Piramila et al. (2012) reported that the physiological stress coursed by the increase in temperature on mungbean (Phaseolus aureus Roxb) and the black gram plant. Global climate change not only impacted by temperature but also depends on CO2 concentration in the atmosphere. Therefore, invasive weeds (Lantana and Parthenium) may become more aggressive under elevated temperature and higher CO2 condition (Naidu, 2014). It is easily understandable from our study that elevated temperatures enhance the growth of E. crus-galli and L. prostrata more than that of M. vaginalis and rice. E. crus-galli showed the increase in height while L. prostrata showed the increase in dry weight and leaf area under elevated temperatures. It was detected that plant height and dry weight of perennial weed species of S. sagittifolia was much higher under elevated températures compare with rice and other perennial weed species.
The results of this study suggest that the growth of rice and weeds was higher under elevated temperature conditions. The present rate of the emission of greenhouse gases gradually increases of the world’s ambient temperature, causing global warming vis a vis climate change. These phenomena will response to plant for adaptation under elevated temperatures and necessary mechanisms in this situation need to be better elucidated for most of the agricultural crops as well as forest and orchard crops and this is worth considering in the Korean Agricultural Transformation Agenda.

