Introduction
Weeds cause great yield losses and increase labour expenses in crop production. Annual weeds usually produce a huge amount of seeds and regrow from these seeds, while perennial weeds can regrow as new plants from the small fragments of underground parts such as roots, rhizomes, and stolons (Schonbeck, 2011).
Currently, weeds are being controlled with synthetic herbicides, physical and mechanical methods, organic herbicides, and bioherbicides (Cai and Gu, 2016). During the last fifty years, heavy reliance on chemical herbicides has increased crop productivity with prominent ecological and economic impacts (Cordeau et al., 2016). After numerous environmental problems due to detrimental impacts of chemical herbicides, researchers investigated alternative biological control systems against weeds by using bioherbicides (Boyette, 2000). Biological agents such as pathogenic and non-pathogenic fungi and bacteria have the potential herbicidal activity to suppress weeds in the crop production system (Saini and Singh, 2018). Generally, biological control agents can be classified into two categories: classical approach and inundative approach (Muller-Scharer et al., 1999).
Classical approach
The classical biological control is based on a natural predator or pathogen of weeds. It requires one or several years to adequately control of weed populations. Also, the degree of weed control is influenced by climatic conditions, geographic locations, and management practices. The pathogen must occupy host-specific and self-sustaining activities. Also, the pathogen can survive and reproduce without attacking any non-target plants after releasing in new environment. Moreover, a life cycle of the pathogen should repeat several times during the growing season to get numerous opportunities to infect the weeds. For the long-term, the pathogen will reach a balanced equilibrium to eradicate the weed populations completely.
Inundative approach
The inundative biological control resembles the classical biological control where the pathogen will be highly specific to the target weed without any harmful effect to non-target plants. The inundative biological control attempts to overwhelm a weed infestation with propagation materials such as bacterial suspensions or fungal spores at high concentrations. Thus, the mass-produced and large numbers of spores from the pathogen are sprayed onto weeds in a manner similar to herbicide application (Harding and Raizada, 2015).
This review focuses on the use of biological weed control agents, specifically, fungi and bacteria. The objectives of this review are to elucidate the factors affecting biological control of weeds in both conventional and alternative management and to assess the prospective strategies of weed control with bioherbicides.
Incentives to use biological weed control
The use of bioherbicides instead of chemical herbicides has offered positive consequences to ecological systems and human health (Harding and Raizada, 2015). The biocontrol agents can degrade rapidly their residual metabolites after application to the natural environment (Li et al., 2003). The cost related to bioherbicides has also been reported that the commercial value of bioherbicides is lower than that of the associated chemical herbicides (Auld and Morin, 1995). The public perception of pesticides is not generally positive. A telephone survey conducted on Canadian consumers where 70% of participants described that they were more satisfied with the foods that were produced by using biological control agents rather than the use of chemical insecticides (McNeil et al., 2010).
Biological agents in weed control can provide the novel modes of action to inhibit even the growth of herbicide-resistant weeds. Moreover, biocontrol agents can be applied in combination with some synthetic herbicides to suppress weeds through the multiple modes of action (Auld and Morin, 1995).
Biotechnology and biological control
The research on biological control faces many challenges, i.e. the screening at a field condition (Harding and Raizada, 2015). In order to create a successful bioherbicide, several complex and specific interactions should be properly conducted between a biological agent and a target weed. An investigation of these interactions in the molecular and biochemical levels helps to find more predictable and efficacious biocontrol agents. These experiments may lead to genetic engineering of highly effective bio-agents which have desired traits (Rosskopf et al., 1999).
It was previously mentioned that transferring a gene from Streptomyces hygroscopicus to Cephalosporium gloeosporioides f. sp. aeschynomene have resulted in a transformant that may infect northern jointvetch (Aeschynomene virginica) as well as the resistant weed, Indian jointvetch (A. indica) (Rosskopf et al., 1999). Similar approaches were used to change the virulence and host range of Xanthomonas campestris pv. campestris with genes encoding the production of bialophos and a range of non-selective phytotoxins produced by S. hygroscopicus (Cobb, 2000).
Phytotoxins produced by phytopathogenic fungi and bacteria
In the last decades, much attention has been paid to several pathogenic fungi and bacteria for their ability to control weeds (Kremer, 2005). Potential biocontrol agents can be isolated from diseased tissues or plants, then, cultured in the selective agar media for their manipulation (Ghosheh, 2005). Plant pathogens interfere with secondary metabolism exudates of the plants by producing a variety of phytotoxins having slightly effects on gene expression of the host up to mortality of the plant (Walton, 1996). The mode of action of phytopathogenic fungi and bacteria can directly interact with specific plant components such as enzyme and membrane receptors. However, if the plant components are absent or altered, no phytotoxic effect is shown on the plant. Thus, molecular targets or phytotoxins are an important determinant of host-pathogen interaction. Phytotoxins can be identified to host-specific phytotoxin and non-specific phytotoxin (Hoagland et al., 2007).
Host-specific phytotoxins
Plant species are affected by host-specific phytotoxins which are produced by pathogenic fungi and bacteria. Some phytotoxins induce toxicity in some host species (Table 1). AAL-toxin of Alternaria alternate is considered as a natural product, since it is one of the biologically derived herbicides. This toxin controls a broad host range of weeds including jimsonweed, duckweed, eastern black nightshade, prickly sida, and hemp sesbania. Most of the host-specific phytotoxins were derived from fungal pathogens (Hoagland et al., 2007).
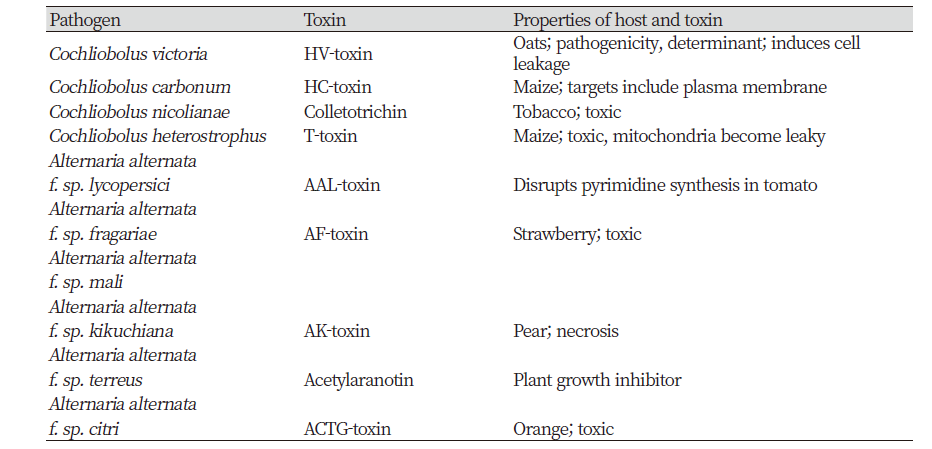
Non-specific phytotoxins
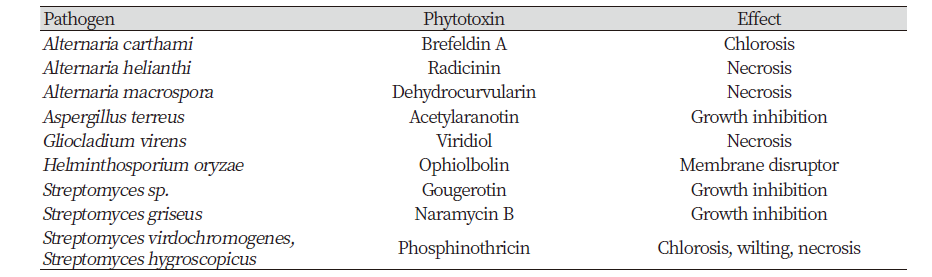
Non-specific phytotoxins are produced by generalist and specialist plant pathogens. The Fusarium genus includes insect pathogens, plant pathogens, and fungal biocontrol species. Chemical contaminants produced by Fusarium such as trichothecenes, fumonisins, naphthazarin, fusaric acid, and related pyridine derivatives induce varying degrees of phytotoxicity to plant tissues and toxicity to mammals (Desjardins, 1992). A causal organism of brown spot disease (Alternaria alternate f. sp. citri) of tangerine orchards can exhibit host-specific and non-specific phytotoxins such as tentoxin and tenuazonic acid from broth culture (Kono et al., 1986). Table 2 shows other selected non-specific phytotoxins by Hoagland et al. (2007).
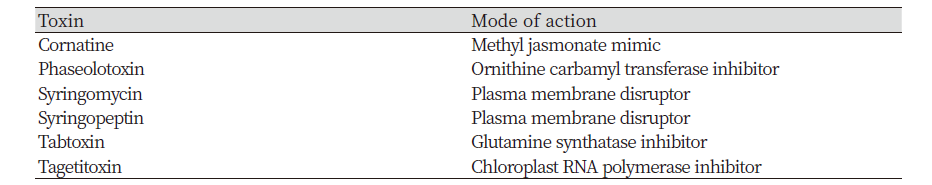
Bacteria induce many diseases in crops and weeds. Among them, Pseudomanas and Xanthomonas were studied as potential biological agents (Harding and Raizada, 2015). Since the discovery of tabtoxin produced by Pseudomonas syringae var. tabaci, other important bacterial compounds have been found in Pseudomonas species. Pseudomonas syringae has been registered as a biocontrol agent to control Canada thistle (Johnson et al., 1996). Table 3 shows other Pseudomonas syringae phytotoxins. Xanthomonas campestris has also been registered to control Poa annua (annual bluegrass) (Bender et al., 1999).
Limitations of biological weed control
The narrow host range, specific requirements of the formulation to assure biocontrol agent, and possible by-products of potent toxins to mammalian by some fungal pathogens limit the development of commercial bioherbicides. Some limitations of biological weed control are as follows.
Increasing host range of bioherbicides
Several pathogens are combined into a single application and then, sprayed on the weeds in pre- and post-emergence applications that are called 'multiple-pathogen strategy' (Charudattan, 2001). For example, the inoculum mixtures including Drechslera gigantia, Exserohilim lingirostratum, and Exserohilim rostratum, which are isolated from different host weeds, completely suppressed the growth of seven weeds of citrus (Chandramohan and Charudattan, 2003). Eight weedy Amaranthus species in soybean were successfully controlled by the conidial suspensions of Phomopsis amananthicola and Microsphaeropsis amaranthi (Otriz-Ribbing and Williams, 2006).
The formulation that improves bioherbicides
Many plant pathogens need definite humidity levels (dew periods) and temperature to be effective during germination, penetration, infection, and killing of the target weeds. This period ranges from 6 hours to more than 24 hours depending upon the interaction of pathogen and the host weed (Boyette, 2000) (Table 4). The important formulations of agrochemicals consist of the forms of emulsions, hydrophilic polymers, organosilicone surfactants, and encapsulated granules, which have to promote virulence and ease of application (Hallett, 2005). The primary functions of these formulations should be emphasized during predisposal of weeds to infection by pathogen and protection of the infected pathogen against environmental impacts while promoting disease development (Charudattan, 2001).

Potential risks of bioherbicides to human health
Some bioherbicides derived from fungal pathogens exhibit strong herbicidal activities to suppress the wide range of harmful weeds, however, these fungal pathogens cause undesirable toxins to mammalian. For example, herbicidal metabolites produced by Myrothecium verrucaria are highly toxic for controlling weeds. Unfortunately, this pathogen simultaneously produces the mammalian toxic i.e. macrocyclic tricothecenes (Anderson and Hallet, 2004). Abbas et al., (2002) revealed that macrocyclic tricothecenes is cytotoxic to mammalian cell lines (half-maximal of proliferation in the concentration range 1-35 nM). Therefore, fungal pathogens that produce both herbicidal compounds and toxins to mammalian should be fully investigated to assess their benefits and risks as potential bioherbicides.
Integrated weed management with biocontrol agents
Integrated weed management is a long-term approach to control weeds through a combination of cultural practices, for instance, herbicides application, genetic technology, allopathy effect, and biological control techniques to minimize the weed seed bank in the soil, prevent weed emergence, and decrease the crop-weed competition (Aldrich and Kremer, 1997). Like chemical herbicides, bioherbicides help to increase the efficacy of weed management in a crop production system.
Management of weed seeds bank in the soil
In the agricultural practice, tillage reduces germination and seedling emergence of weeds in the upper layer of soil, and which allows doing surface applications of bioherbicide to close proximity with weed seeds (Clements et al., 1996). The persistence of weed seeds in the soil was reduced by the direct application of bioherbicides with target agents in seed-germination zone and combination with solarisation enhances seeds deterioration in soil (Wagner and Mitschunas, 2008). The isolation of Epicoccum purpurascens Ehrenb from Echinochloa spp. inhibited seed germination of Echinochloa in the rice field (Motlagh, 2011). Ahmed et al. (2001) identified Fusarium solani which reduced the seed germination of Striga significantly.
Mechanical weeding
Weeds can be controlled effectively by mechanical weeding at cotyledon and seedling stages. A period for weed control should overlap with the implementation of mechanical weeding and bioherbicide application processes.
Suppression by crop cultivars
The selection of crop cultivars is essential in completing integrated weed control. Bioherbicides can be used as an additional tool to decrease the growth of weeds and increase crop-weed competition (Cordeau et al., 2016).
Management of herbicide-resistant weeds
Integrated weed management systems substantially reduce the populations of herbicide-resistant weeds in the field (Oven et al., 2015). Currently, the resistance of weeds to herbicide is a serious problem in crop production and is encouraged by the increasing dependence on a limited number of active ingredients. Therefore, bioherbicides should be considered as an alternative control strategy to minimize the selection pressure of weeds, whereas chemical herbicides are usually not effective to control these weeds due to the evolved resistance mechanisms.
Conclusion
Weeds can be controlled by chemical herbicides to prevent invading new areas as a short-term solution. However, the use of board-spectrum chemical herbicides has resulted in a reduction to the biodiversity of natural enemies, an outbreak of weeds, development of herbicide-resistant weeds, and contamination of food and ecosystem. For long-term control, biological agents should be integrated into weed management systems to minimize weed problems in sustainable agriculture and to develop sustainable ecosystems.
This review demonstrated that plant pathogens have an expanding role in weed control. There are many products composed of living bacteria and fungi that claim they will increase plant health. However, biological control with plant pathogens in weed control should be considered as an ecological safe when compared to other established practices of weed control. To have a sustainable agricultural system, a combination of all available methods should be integrated into weed management to minimize contamination and risks to the environment.
Acknowledgements
This work was carried out with the support of the “Cooperative Research Program for Agriculture Science & Technology Development (Project No. PJ01347901)”, Rural Development Administration, Republic of Korea.
Authors Information
Kee Woong Park, Department of Crop Science, Chungnam National University, Professor
Kwang Min Cho, Department of Crop Science, Chungnam National University, Researcher
Jung Sup Choi, Korea Research Institute of Chemical Technology, Researcher
Botir Khaitov, Department of Crop Science, Chungnam National University, Researcher
Aung Bo Bo, Department of Crop Science, Chungnam National University, Ph.D. student, https://orcid.org/0000-0001-7579-3429
Mirjalol Umurzokov, Department of Crop Science, Chungnam National University, Ph.D. student

