서언
전 세계적으로 국가 간 인적, 물적 교류가 늘어남에 따라 외래식물의 유입 경로가 다양화되고 있으며, 이에 따라 외래식물의 국내 유입이 증가하고 있다(Son et al., 2019; Son et al., 2021). 외래식물은 정착 시 자생종의 생육지 파괴, 생태계 교란, 생물다양성 감소, 작물 수확량 감소 등 여러 측면에서 부정적인 영향을 미치는 것으로 알려져 있다(Son et al., 2021; Higgins et al., 1999; Pimentel, 2005; Gaertner et al., 2009; Jung et al., 2017). 유입된 외래식물을 통해 발생할 수 있는 위해성으로부터 생태계를 보호하기 위해서는 이들의 현황, 유입 경로 파악, 생태계에 미치는 영향 파악 등 관리하는 것이 필요하다(Kang et al., 2020; Kil and Kim, 2014). 국내의 경우 유입된 외래생물 중 위해성평가 결과 생태계 등에 미치는 위해가 큰 것으로 판단된 17종을 생태계교란 식물로 지정하여 관리하고 있다(KLIC, 2023).
미국쑥부쟁이(Aster pilosus, Syn. Symphyotrichum pilosum)는 북아메리카 원산의 국화과(Asteraceae) 다년생 초본으로 하천, 시설지, 나지, 도로 주변 등 광역적으로 분포하고 있다(Chmielewski and Semple, 2001; Kil and Kim, 2014; Oh et al., 2011). 미국, 캐나다, 유럽(프랑스, 독일, 스페인, 벨기에, 이탈리아 등), 아시아(인도, 한국 등)에 서식하고 있는 것으로 확인되었다(Muligan, 1992; Park et al. 2021; Fehér, 2008). 국내에 서식하는 미국쑥부쟁이는 원예용 식물 또는 군수품 도입 시 유입된 것으로 추정되며, 1970년대 말 강원도 춘천시 중도에서 처음 발견되었다(Kim et al., 2008). 현재 충북 및 경북의 일부 지역과 경기도, 강원도 지역에 집중적으로 분포하고 있으며, 경기도와 강원도에서 남쪽으로 확산되고 있다(Kim el al., 2017). 유입된 미국쑥부쟁이는 정착한 지역의 식생을 단순화시키며, 종 다양성과 자생식물의 생육을 감소시킨다(Yoon et al., 2022). 이로 인해 유럽과 호주의 여러 지역은 미국쑥부쟁이를 문제를 일으키는 잡초로 간주하고 있으며, 미국에서는 “Composite List of Weeds”에 등재하여 관리하고 있다(Chmielewski and Semple, 2001; Rhoads and Block 2007). 국내의 경우 생태계에 교란을 야기할 우려가 높아 환경부에서 2009년 생태계교란 식물로 지정 고시하여 관리하고 있다(Yoon et al., 2022).
현재 미국쑥부쟁이의 확산은 빠르게 진행되고 있으며, 높은 출현빈도로 향후 전국에 출현할 가능성이 높은 종으로 서식특성을 반영한 맞춤형 사업방식 및 반복제거, 대체수종 식재 등 지속적인 관리가 필요하다(Park and Lee, 2018; Shin et al., 2017). 외래식물을 효과적으로 관리하기 위해서는 각 외래식물에 대한 생육 특성을 파악하고 이들의 확산 요인을 분석하는 것이 필요하며, 종의 서식 범위 내에서 발아조건과 출현패턴의 차이를 이해하는 것이 중요하나 국내에 서식하고 있는 미국쑥부쟁이에 대한 발아특성 연구는 미비한 실정이다(Son et al., 2021; Blossey et al., 2017).
따라서, 본 연구는 확산 우려가 큰 미국쑥부쟁이를 대상으로 온도 및 광조건에 따른 발아반응을 조사함으로써 외래식물의 확산 방지 및 효율적인 방제를 위한 기초자료를 제공하고자 수행하였다.
재료 및 방법
실험재료
연구에 사용된 미국쑥부쟁이(Aster pilosus) 종자는 2023년 1월 3일 충청북도 청주시 미호천(36°44'05.8"N 127° 29'01.6"E)에서 채종하였다. 채종한 종자는 정선과정을 거친 후 실험에 사용하기 전까지 밀봉하여 상온에 보관하였다.
기본특성 조사
종자의 외부 형태는 국립백두대간수목원 공동기기센터의 광디지털현미경(DVM6, Lieca)을 이용하여 촬영하였으며, 길이와 너비는 무작위로 선발한 종자 25립을 측정한 후 평균과 표준오차를 구하였다. 종자의 천립중은 전자저울로 1,000의 무게(mg)를 4회 반복 측정하여 평균과 표준오차를 구하였다.
온도와 광조건에 따른 발아반응
종자는 치상 전 1% NaClO 용액에 2분간 침지 소독한 후 증류수로 5회 이상 수세하였다. 모든 처리구는 petridish (90 ×15 mm)에 여과지(90 mm, No. 2, Advantec, Toyo) 2매를 깔고 증류수를 충분히 주입한 후 25립 4반복으로 치상하였으며, 수분 손실 방지를 위해 실링테이프로 밀봉하였다. 온도에 따른 발아반응을 알아보기 위해 항온(10, 15, 20, 25, 30, 35℃)과 변온(15/6, 20/10, 25/15, 30/15, 35/20℃)으로 완전임의 배치하여 배양하였다. 광조건에 따른 발아반응을 알아보기 위해 30, 30/15℃의 온도조건에서 명조건(12/12h, L/D)과 암조건(24h, L/D)으로 구분하여 진행하였으며, 광도는 약 14 μmol·m-2·s-1으로 조사하였다. 암조건의 경우 명조건과 동일한 조건하에 알루미늄 지퍼백을 이용하여 광을 차단하였으며, 30일 이후 최종발아율을 확인하였다.
발아특성 조사
치상된 종자는 2일 간격으로 30일 동안 발아율을 조사하였으며, 발아는 유근이 1 mm 이상 출현하였을 때를 기준으로 하였다. 조사 기간 중 발아된 종자는 제거하였으며, 조사 시 부패한 종자는 발아특성 계산에서 제외하였다. 각 처리별 발아 조사 결과를 이용하여 발아율 (Germination rate, GR), 발아개시일 (Frist day of germination, FDG), T50 (Days to 50% of germination), 평균 발아일수 (Mean germination time, MGT), 발아속도 (Germination speed, GS)를 다음과 같은 공식을 이용하여 구하였다(Kader, 2005; Kim et al., 2021).
• GR (%) = (N / S) × 100
• FDG (days) = Day on which the first germination event occurred
• T50 (days) = Ti + (Tj - Ti) × (N/2 - Ni)/(Nj - Ni)
• MGT (days) = Σ (ti × ni)/ Σ ni
• GS = ∑(Nx/Tx)
(N; 총 발아수, S; 총 공시 종자수, ti; 치상 후 조사일수, ni; 조사 당일의 발아수, Ni: N에 대한 50% 발아 직전까지의 총 발아수, Nj: N에 대한 50% 발아 직후까지의 총 발아수, Ti: Ni 시점까지 소요된 발아기간, Tj: Nj 시점까지 소요된 발아기간, Nx: 조사 당일의 발아수, Tx: 치상 후 조사일수)
통계처리
처리별 발아반응 결과에 대하여 조사된 데이터는 SAS 9.4 (SAS Institute Inc. Cary, USA)를 이용하여 분산분석(ANOVA) 하였으며, 처리의 평균 간 통계적인 유의성은 Duncan's Multiple Range Test(p <0.05)를 이용하여 비교하였다.
결과 및 고찰
기본특성 조사
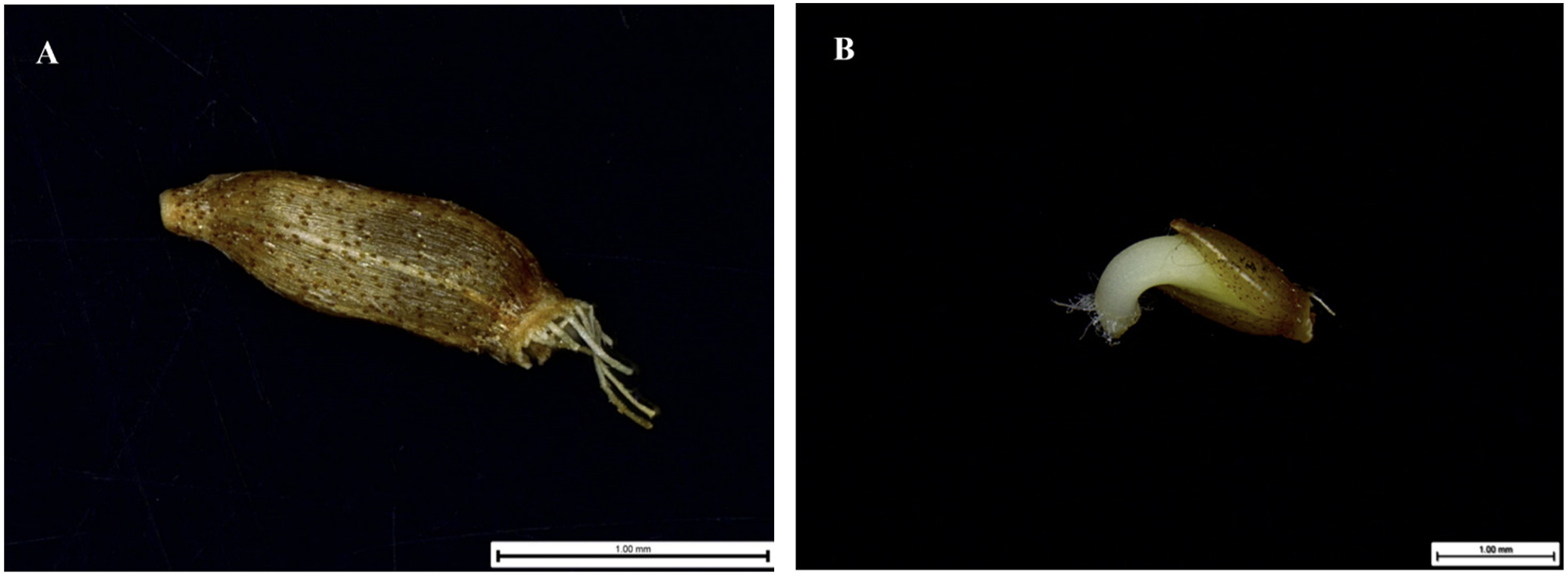
미국쑥부쟁이 종자는 좁은 도란형-원통형(Narrow obovoid/obovate-Cylindrical)에 백색의 관모가 붙어 있는 형태이며, 종피색깔은 연한 갈색에 진한 갈색 점들이 관찰되었다(Fig. 1A). Martin (1946)에 의하면 종자의 길이가 0.3-2.0 mm인 경우 미세종자로 분류되는데 미국쑥부쟁이 종자의 길이와 너비의 길이는 각각 1.43 ± 0.14 mm, 0.53 ± 0.05 mm로 측정되었으며, 천립중은 180 ± 4 mg으로 미세종자로 확인되었다(Table 1). 발아는 종피가 열개된 후 유근이 돌출하면서 이루어졌다(Fig.1B).

온도에 따른 발아반응
명조건에서 온도에 따른 발아반응은 모든 요소에서 유의미한 차이를 보였다(Table 2). 10℃ 조건은 치상 후 18.5일 후 발아가 시작되어 21.5일 후 8.78 ± 3.25%로 가장 낮은 발아율을 보였다. 30℃와 25/15℃ 조건은 6일 만에 발아하기 시작하였으며, 각각 12.5일 후 88.59 ± 4.88%, 20.5일 후 88.63 ± 2.75%로 높은 발아율을 보였다(Fig. 2). 종자의 발아율은 특정 온도범위 내에서 일정하게 유지되고 범위를 벗어나게 되면 감소되며(Thompson, 1970), 미국쑥부쟁이의 경우 온도가 높아짐에 따라 발아율이 증가하지만 30℃, 25/15℃ 이상의 온도에서 감소되었다(Fig. 2).
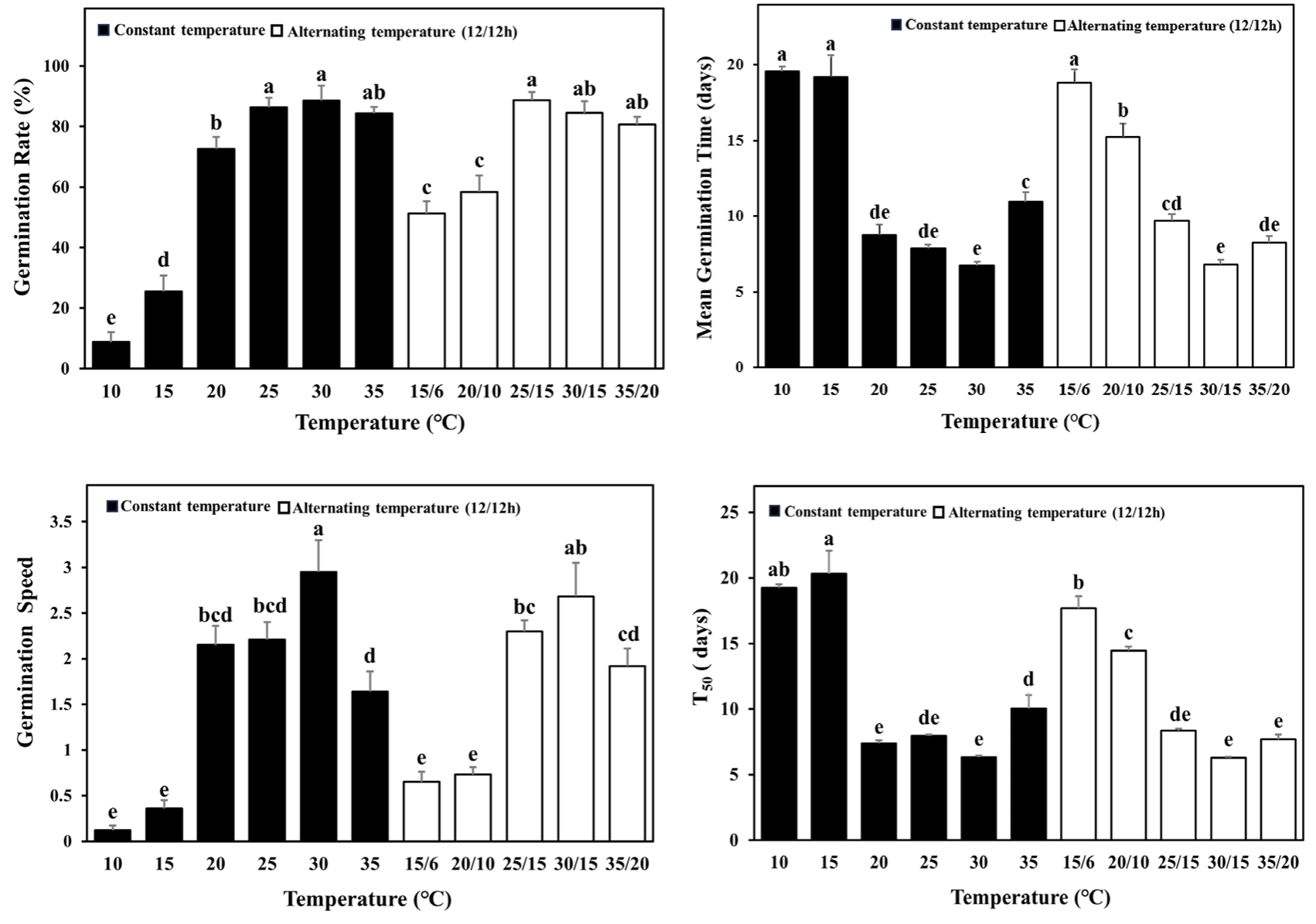
Baskin and Baskin (1985)의 보고에 따르면, 미국쑥부쟁이 종자의 발아율은 15/6℃에서 60%이며, 20/10℃ 이상의 온도에서 98-99%로 본 연구와 유사한 결과를 보였다. Baskin and Baskin (1979)의 경우 15/6℃에서 10%의 발아율을 보여 53.1%의 발아율을 보인 본 연구와 차이를 보였으나 온도가 증가함에 따라 발아율이 증가하다 30/15℃ 이상의 온도에서 감소하는 경향은 유사하였다.
종자의 발아는 채종 환경과 저장 온도 등에 따라 다양하게 나타날 수 있으며(Han et al., 2022), 미국쑥부쟁이의 경우 조도(Bazzaz 1968; Baskin and Baskin, 1979, 1985), 토양연령(Jackson and Willemen, 1976), 수과의 무게(Prinzie and Chmielewski, 1994) 등의 영향을 받는다. 따라서, 이러한 차이는 채종환경, 조도 등 실험에 사용된 미국쑥부쟁이 종자와 실험조건의 영향으로 발생한 것으로 사료된다.
미국쑥부쟁이의 T50은 전체적으로 6-20일로 조사되었다. 그 중 15℃는 20.32 ± 1.74일로 가장 길었으며, 30℃와 30/15℃는 각각 6.32 ± 0.12일, 6.29 ± 0.08일로 가장 짧은 것으로 조사되었다. 평균발아일수는 10℃에서 19.55 ± 0.33일로 가장 길었으며, 30℃와 30/15℃에서 각각 6.73 ± 0.26, 6.81 ± 0.31일로 조사되었다. 이는 미국쑥부쟁이의 발아는 5-7일에 최고점에 도달한 후 점진적으로 감소된다는 Prinzie and Chmielewski (1994)의 보고와 유사한 결과를 보였다. 발아속도는 10℃에서 0.12 ± 0.05로 가장 느렸으며, 30℃와 30/15℃에서 각각 2.95 ± 0.35, 2.68 ± 0.37로 온도가 높아짐에 따라 발아속도가 증가하는 결과를 보였다. 온도가 높아질수록 T50와 평균발아일수는 감소하였으며, 발아속도는 증가하였다. 30℃와 30/15℃ 이상의 온도에서 T50와 평균발아일수는 길어지고 발아속도는 감소하는 경향을 보였다(Fig. 2 and Table 2). 이는 발아상의 온도가 높아짐에 따라 빠르게 발아가 이루어졌으나, 30℃와 30/15℃ 이상의 온도에서 발아율이 감소하였기 때문으로 판단된다.
광조건에 따른 발아특성
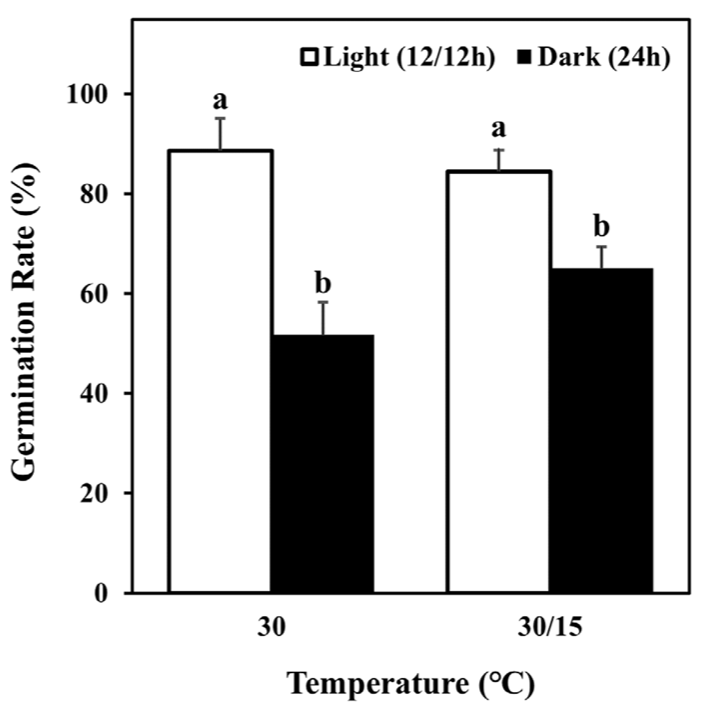
미국쑥부쟁이는 광조건에 따른 발아율에 유의미한 차이를 보였다(Fig. 3). 명조건의 경우 30℃와 30/15℃에서 각각 88.59 ± 4.88, 84.45 ± 3.85%의 발아율의 보였으나, 암조건에서 51.73 ± 5.2, 65.09 ± 4.33%로 명조건에 비해 낮은 발아율을 보였다(Fig. 3). 이는 암조건의 모든 온도조건에서 0-22%의 발아율을 보인 기존 연구와 차이를 보였으나, 암조건보다 명조건에서 높은 발아율을 보이는 경향은 본 연구의 결과와 일치하였다(Baskin and Baskin, 1979; Baskin and Baskin, 1985).
미국쑥부쟁이 종자는 겨울까지 분산되지 않을 경우 식물체에 있는 동안 성숙되며, 층적 요구 상황을 충족한다(Baskin and Baskin, 1985). 층적기간 동안 광에 노출되면 발아에 필요한 광 요구량을 충족시킬 수 있으며, 광 요구량이 충족되면 암조건에서 발아한다(Baskin and Baskin, 1979; Baskin and Baskin, 1985). 겨울에 1-10주 동안 광을 받은 미국쑥부쟁이의 경우 암조건에서 2-67%의 발아율을 보인다(Baskin and Baskin, 1985). 따라서, 이러한 차이는 실험에 사용된 종자가 채종 전에 발아에 필요한 광 요구량을 충족하여 발생한 차이로 사료된다.
발아반응을 종합적으로 고려했을 때, 항온조건의20-35℃와 변온조건의15/6-35/20℃가 미국쑥부쟁이가 발아할 수있는 온도 범위이며, 그 중 30℃와 30/15℃ 조건에서 높은 발아율을 보일 것으로 판단된다. 이에 따라, 국내의 최고기온과 최저기온을 고려하였을 때 미국쑥부쟁이 종자는 3월 초부터 발아하기 시작할 것으로 예상된다. 이는 미국의 노스캐롤라이나주 (North Carolina)에서 3월 이전에 확인할 수 있으며, 켄터키주(Kentucky)에서는 3월초에 미국쑥부쟁이를 확인할 수 있다는 보고와 일치한다(Baskin and Baskin, 1979; Keever, 1950). 따라서, 종자의 성숙시기 얕은 경운과 3월 중순 손제초를 통해 미국쑥부쟁이를 방제할 수 있을 것으로 보이며, 본 연구에서 도출된 온도와 광에 따른 미국쑥부쟁이의 발아반응은 확산 방지 및 효율적인 방제를 위한 대책 수립 과정에 기초자료로 활용될 수 있을 것으로 판단된다. 하지만 본 연구는 특정 서식지에서 특정 연도에 수집한 종자를 사용하여 수행되었기 때문에 다양한 서식지와 시기에서의 종자 수집이 고려되지 않았다는 한계가 있다. 따라서, 향후 연구에서는 다양한 서식지와 시기에서 채종한 종자를 활용하여 실험을 진행함으로써 환경에 따른 영향을 파악할 필요성이 있다.
요약
미국쑥부쟁이(Aster pilosu)는 국화과 다년생잡초로 대한민국에서 생태계교란종으로 지정되어 있다. 본 연구는 생태계교란 식물 미국쑥부쟁이 종자의 발아특성을 조사하기 위해 진행되었다. 2023년 1월 3일 청주시에서 채집한 종자를 실험에 사용하였다. 발아검정은 항온조건 (10, 15, 20, 25, 30, 35℃)과 변온조건 (15/6, 20/10, 25/15, 30/15, 35/20℃)에서 30일 동안 수행하였다. 종자의 외부 형태를 관찰한 결과, 너비와 길이는 각각 0.53 mm, 1.43 mm로 측정되었으며, 천립중은 180 mg으로 조사되었다. 미국쑥부쟁이는 항온 25-35℃와 변온 15/6-35/20℃에서 80-88%의 발아율을 보였으며, 10℃와 15℃에서 각각 8%, 25%로 낮은 발아율을 보였다. 광조건에 따른 발아검정 결과, 미국쑥부쟁이 종자는 암조건보다 명조건에서 발아율이 높았으며, 암조건에서 65% 이하의 발아율을 보였다. 따라서, 항온조건의 20, 25, 30, 35℃와 변온조건의 15/6, 20/10, 25/15, 30/15, 35/20℃가 미국쑥부쟁이가 발아할 수 있는 온도범위이며, 그 중 명조건의 30℃와 30/15℃조건에서 높은 발아울을 보일 것으로 판단된다.

