서 언
농업인구의 감소에 따라 노동력 감소를 위한 생력재배기술의 개발이 요구되고 있다. 실제 2014년 국내 농업인구는 1990년에 비해 58% 감소하였고 특히 농업인 비율 중 60대 이상의 비율이 39%을 차지하여 농업 노동력의 감소가 큰 것을 알 수 있다(Korean Statistical Information Service, 2014). 벼 직파재배방법은 노동력 절감의 대표적인 기술이며 벼 직파재배방법에는 건답직파, 무논직파 재배방법이 개발되어 있다(Back et al., 1998; Kim et al., 1991; Kim et al., 1995). 벼 직파재배방법은 이앙재배방법과 비교하여 육묘 및 이앙작업 생략에 따른 노동력 절
감 효과는 가지고 있지만 초기 입모의 불안정, 도복의 위험성 등 몇 가지 단점도 가지고 있다. 또한 직파재배는 이앙재배에 비해 잡초 및 잡초성벼의 발생이 많으며, 이들 잡초 및 잡초성벼 발생은 벼의 수량을 크게 감소시킨다고 알려져 있다(Kim, 1997). 특히 잡초성벼는 벼와 유전적인 조성이 같아 벼와 동시에 포장에 발생 시 화학적 방제가 불가능해 직파재배지역에서 큰 문제점이 되고 있다(Smith, 1989; Akasaka et al., 2009; Londo and Schaal, 2007). 1995년 1,175,000 ha까지 증가한 벼 직파재배면적은 이와 같은 재배상의 어려움으로 2014년 178,000 ha까지 감소하였다.
잡초성벼는 재배벼에 비해 낮은 온도에서도 발아력이 높고 다양한 환경스트레스에도 강한 특성이 있어 재배벼와 경쟁하여 생육할 수 있는 여러 가지 이점을 가지고 있다(Hoagland and Paul, 1978; Cho et al., 2013; Chung and Paek, 2003; Kim et al., 2009). 벼 직파재배의 안정적인 확립을 위해 잡초성벼 방제의 중요성이 확대되면서 잡초성벼의 발아 및 출아특성 분석을 위한 시험은 다수 진행되었으나 몇 점의 특정 잡초성벼 유전자원을 대상으로 한 실험들이 주를 이루고 있다(Cho et al., 2013. Im et al., 2004). 하지만 현재 잡초성벼의 다양성이 증가하고 있으며 실제 유전자원 센터에는 국내에서 수집된 잡초성벼 5,300여 점이 등록되어 있어 적은 수의 유전자원을 대상으로 한 실험 결과는 잡초성벼의 특성을 대표적으로 나타내기에는 어려움이 있다고 사료된다. 이에 본 실험은 잡초성벼의 효율적인 방제를 위한 기초자료를 제공하기 위해 다양한 유전자원을 이용하여 잡초성벼의 출아특성을 분석하고자 하였다.
재료 및 방법
국내 수집 잡초성벼 유전자원 구분 및 시험재료 선정
국내 수집된 잡초성벼 유전자원 중 시험재료를 선정하기 위해 유전자원센터에 등록된 잡초성벼 유전자원 5,300여점을 대상으로 수집지역을 분석하여 9개 도로 분류하였다(Table 1). 수집지역을 분석한 결과 충청남도와 경상북도에서 각 15.5%, 13.5%의 유전자원이 수집되어 가장 많은 잡초성벼 유전자원이 수집, 등록된 것으로 나타났다. 잡초성벼 유전자원의 수집지역을 분류하여 각 지역의 분포 정도를 파악 후 분포 정도에 맞게 유전자원 개수를 배분하여 총 100점의 유전자원을 무작위로 시험재료로 선정하였으며, 수집 유전자원 중 등록된 유전자원 명이 다양하여 다양성이 인정되는 지역은 시험재료를 더 추가하여 선정하였다. 그 결과 충청남도 수집 유전자원 19점, 경상남도 및 전라북도에서 14점, 경상북도 및 전라남도 13점, 충청북도 6점, 제주도 5점, 강원도 수집 유전자원 중 7점을 시험재료로 선정하였으며 이들을 유전자원센터에서 분양 받아 사용하였다(Table 1).
휴면정도에 따른 시험재료 분류
분양 받은 잡초성벼 유전자원의 종피색, 현미 장폭비, 천립중 등 일반 특성을 조사한 뒤 이를 증식하여 수확 후 각 유전자원의 휴면성을 분석하였다. 패트리디쉬에 watman #2 여과지를 깔고 각 시험재료를 30립씩 3반복으로 치상한 뒤 증류수를 5 ml 첨가하여 발아시험을 수행하였다. 발아시험 온도조건은 30°C 항온조건으로 설정하였고 암 발아성 및 광 발아성 검정을 위해 은박지를 씌운 처리와 무처리로 발아시험을 진행하였으며, 발아 치상 3주 후 유근이 1 mm 이상 나타난 것을 발아한 것으로 조사하였으며, 발아하지 않는 종자를 휴면성을 가지고 있는 것으로 판단하였다. 발아 조사 후 발아율이 80% 이상으로 일반 재배벼와 비슷한 수준을 보이는 유전자원을 휴면성이 약한 것으로, 발아율이 80% 이하인 것을 일반 재배품종에 비해 휴면성이 높은 것으로 분류하여 시험재료로 사용하였다.
생육온도 및 토양매몰깊이에 따른 출아율변화 분석
수확 후 발아율 검정 결과 발아율이 일반 재배품종와 비슷한 수준인 80% 이상으로 나온 시험재료를 이용하여 생육온도 및 파종심도에 따른 출아변화를 검정하였다. 남아있는 휴면성을 소거하고, 수분 흡수율에 따른 출아율 오차를 제거하기 위해 종자를 0.1% 질산용액에 2일간 처리하였으며 수온은 10°C 밑으로 유지하여 발아가 되지 않도록 하였다. 생육실험은 식물생장상(Conviron, PGC-20)을 이용하였으며, 생육온도는 4처리로 하여 주/야간 온도를 15/5°C, 16/6°C, 17/7°C, 18/8°C, 19/9°C (평균온도 각 10, 11, 12, 13, 14°C)로 설정 하였으며 일장은 13시간으로 설정하였다. 파종심도는 1, 3, 5 cm로 설정하였고 흙은 일반 논 포장의 토양을 채취하여 건조 후 2 mm 체를 통과한 흙을 사용하였다. 트레이에 흙을 1 cm 깊이로 깔고 6×6×8 cm로 제작된 플라스틱 칸막이를 꼽고 각 시험재료를 30립씩 3반복으로 파종 후 흙을 1, 3, 5 cm로 덮었다. 물은 아래에서 공급하여 흙에 스며들도록 처리하였으며 토양의 건조를 막기 위해 4일에 한번씩 물을 공급하였다. 흙 위로 식물체가 1 cm 이상 나왔을 때를 출아한 것으로 정하여 출아 정도를 조사하였다(Fig. 1A).
동계 야외처리에 의한 출아율변화 분석
수확 후 발아율 검정 결과 발아율이 80% 이하인 시험재료는 일반 재배품종에 비해 휴면성이 높은 것으로 판단하였다. 이들 유전자원은 벼 수확기 때 탈립 후 바로 출아하지 않고 휴면성이 소거된 뒤 출아가 나타나게 된다. 실제 포장에서와 같이 휴면성 소거 뒤 출아율 변화를 분석하기 위해 이들 유전자원은 수확 후 휴면성을 유지한 채 파종을 하여 겨울 동안 야외처리를 하였으며, 이듬해 봄 출아율 검정을 실시하였다. 트레이에 흙을 1 cm로 높이로 깔고 6×6×11 cm의 플라스틱 칸막이를 끼운 뒤 휴면성이 유지된 각 시험종자를 30립씩 3반복으로 파종하였다. 파종심도는 1, 3, 5, 7 cm로 설정하여 흙을 덮은 뒤 물을 주고 야외에 처리하였다(Fig. 1B). 야외처리는 2015년 11월 1일에 시작하였으며 겨울 동안에는 인위적인 물 공급을 하지 않고 2016년 3월부터 출아유도를 위해 물 공급을 시작하였다. 4월부터 출아가 시작되어 조사를 시작하였다.
Fig. 1.
Cultivation of weedy rice for testing of emergence characters using (A) low dormancy and (B) high dormancy weedy rice genetic resources. Low or high dormancy weedy rice was classified according to the germination rate of 80%. Seed was sowed at different soil depth conditions with three replications. Low dormancy weedy rice was cultivated at 15/5°C, 16/6°C, 17/7°C, 18/8°C, 19/9°C with 13 hours light condition. High dormancy weedy rice was cultivated at natural condition from November in to 2015 to May in 2016.

통계 및 데이터 분석
생육온도에 및 매몰 깊이에 따른 출아변화 분석은 SAS Version 9.2 (SAS Inc. US) 를 이용하여 Duncan’s multiple test로 p<0.05 수준에서 유의적 차이를 검정하였다. 출아까지 영향을 미치는 적산온도를 계산하기 위해 파종부터 출아까지 일 평균온도에서 출아가 진행되지 않는 온도인 10°C를 제한 일 평균 온도를 합산하여 유효 적산온도를 계산하였다. 유효적산온도에 따른 잡초성벼의 출아 정도를 분석을 위해 매몰 깊이 별 최종발아율을 100%로 두고 각 조사시점의 발아율을 상대발아율로 계산하였으며, 유효적산온도에 따른 상대발아율변화는 아래와 같은 logistic equation을 사용하여 계산하였다(Verhulst, 1838).

F는 출아율, A는 최종 출아율, RF는 최대 출아속도, t는 유효적산온도, tm은 최종 출아율이 절반이 되는 유효적산온도 각 의미한다. A, RF, t, tm 값은 비선형 회귀분석에 의해 결정되는 계수이다.
결과 및 고찰
잡초성벼 유전자원 일반특성 및 휴면정도에 따른 분류
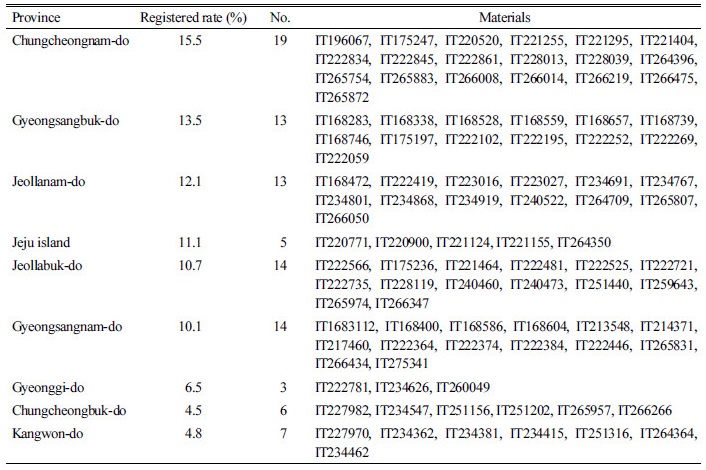
유전자원센터로부터 분양 받은 국내 수집 잡초성벼 유전자원 100점의 특성조사를 실시하였다. 종피색에 따라 분류한 결과 왕겨 및 종피색이 붉은색인 적미는 각각 80, 79점, 왕겨 및 종피색이 백색인 유전자원은 20, 21점으로 적미의 비율이 큰 경향이었다(Fig. 2A). 현미 장폭비 분석 결과 1.96-3.58였으며 장폭비가 2.2-2.59 사이인 유전자원의 비율이 60%로 장형을 나타내는 유전자원이 많은 것으로 나타났다(Fig. 2B). 현미 천립중은 12.8-28.9g이었으며, 유전자원의 72%가 일반 재배벼와 비슷한 수준인 17-21.9g의 현미 천립중을 가진 것으로 나타났다. 현미 색에 따른 천립중 분석결과, 현미 천립중이 23g 이상인 대립종은 백미인 것으로 나타났다(Fig. 2C). 유전자원 100점을 증식 후 2일간 자연광에서 건조한 종자의 명조건에 따른 발아율을 조사한 결과 암 발아율은 명 발아율의 0.94배로 증가 하였다(Fig. 2D). 평균 발아율은 49%로 발아율이 80% 이상인 유전자원이 40점, 80% 이하인 유전자원이 60점 인 것으로 나타났으며 현미색에 따른 발아율의 차이는 보이지 않았다(Fig. 3).
휴면성 낮은 유전자원의 출아특성 분석
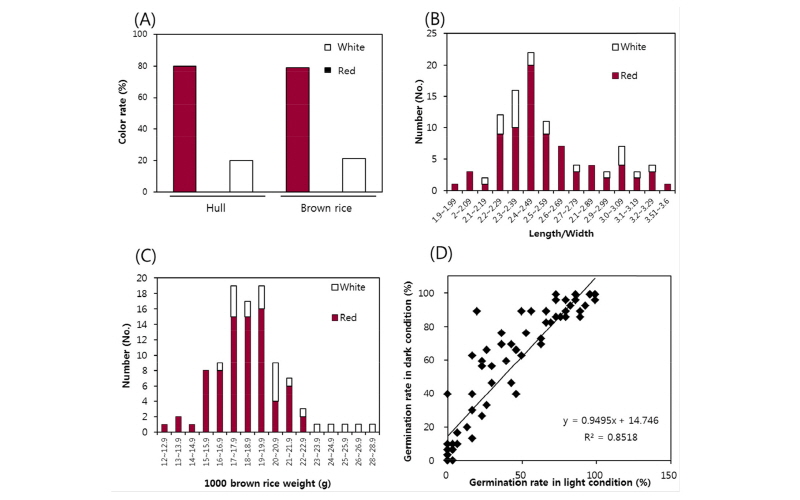
발아율이 일반 재배품종와 비슷한 수준인 80% 이상으로 휴면성이 낮은 유전자원 40점을 대상으로 생육온도 및 파종심도에 따른 출아특성을 분석하였다(Fig. 3). 시험재료 40점 중 적미는 31점, 백미는 9점이었다. 생육 평균온도를 10, 11, 12, 13, 14°C에서 처리 후 30일까지의 출아특성을 분석한 결과 평균온도 10°C에서는 파종 후 30일까지 출아가 나타나지 않았다. 파종심도 1 cm의 경우, 11°C에 비해 12°C에서 출아율이 적미의 경우 27%, 백미의 경우 40% 상승하였으며, 12°C와 13°C에서는 큰 사이를 보이지 않았다. 반면 14°C에서는 적미와 백미의 출아율이 13°C에 비해 65% 및 57% 상승하는 것으로 나타났다. 이를 통해 잡초성벼 유전자원은 평균온도 11°C 이상에서 출아가 가능하며 출아에 큰 영향을 미치는 온도는 12°C 및 14°C로, 평균온도 12°C를 넘어야 출아가 활발히 시작되며 왕성한 출아율이 나타나기 위해서는 평균 14°C이상이 되어야 할 것으로 사료된다. 평균온도가 11°C로 낮을 경우 종피색에 따른 출아율 차이가 뚜렷하게 나타났는데 이는 적미가 저온에서 발아율이 높다는 기존의 보고들과 같은 결과로 판단된다(Suh et al., 1992; Yuan et al., 2007). 파종심도에 따른 출아율 분석 결과, 파종심도가 깊어질수록 출아율이 감소하는 경향이었으며, 파종심도에 따른 출아율 감소는 생육온도가 낮아질수록 유의적으로 감소하였다. 하지만 적미의 경우 백미에 비해 파종심도에 따른 출아율 감소가 적은 것으로 나타났는데 이는 앵미의 발아특성 분석결과 파종심도가 6 cm로 깊어져도 일반품종에 비해 발아율이 높다는 기존에 보고와 유사한 결과로 사료된다(Cho et al., 2013).
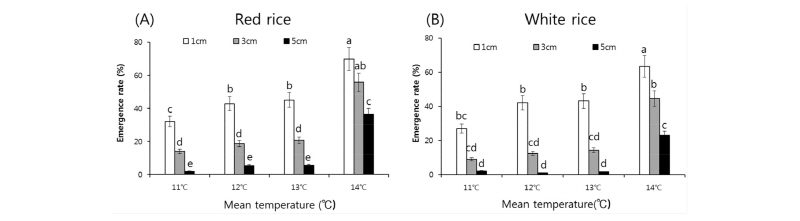
각 생육온도의 파종심도에 따른 출아율 데이터를 이용하여 유효적산온도에 따른 출아율의 변화를 분석하였다(Fig. 4). 파종심도가 깊어질수록 적산온도와 출아율의 유의관계는 감소하였지만 파종심도 1 cm 및 3 cm에서는 상관계수 각각 0.76, 0.70으로 적산온도에 따른 출아율 변화에 상당한 유의성이 나타났다. 평균온도가 11°C 이상일 시 온도가 1°C 올라갈수록 파종심도 1 cm에서는 출아율이 0.6%, 3 cm에서는 0.5% 증가하였으나, 5 cm에서는 0.28% 증가하는 경향으로 파종심도가 5 cm이상이 될 시 온도증가에 따른 출아율 상승률이 급격히 감소되는 경향이었다.
휴면성 높은 유전자원의 출아특성 분석
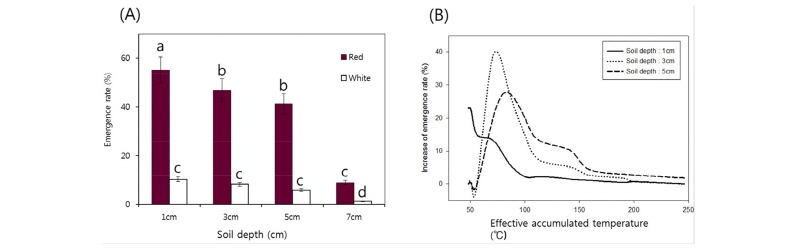
시험재료 중 수확 후 발아율 검정결과 발아율이 80% 미만으로 일반 재배품종에 비해 휴면성이 높다고 판단되는 잡초성벼 유전자원은 60점이었으며 이 중 적미는 49점, 백미는 11점이었다. 이들 유전자원은 겨울철 휴면성이 소거된 뒤 봄철 온도상승에 따라 출아될 것으로 판단되어 동계 야외처리에 의한 이듬해 출아율 변화를 관찰하였다. 파종심도에 따른 출아율 변화를 관찰하기 위하여 파종심도를 1, 3, 5, 7 cm 설정하여 출아율을 분석한 결과는 그림 5과 같다. 더 이상의 출아가 나타나지 않아 출아가 완료되었다고 판단된 5월 중순인 5월 17일 파종심도에 따른 출아율을 분석한 결과(Fig. 5A), 현미색에 따른 큰 차이가 관찰되었다. 적미의 경우 파종심도 1 cm, 3 cm, 5 cm에서 출아율이 55%, 46%, 41%로 다소 높았고 파종심도 7 cm에서는 출아율이 8.8%로 크게 감소하였다. 반면 백미의 경우 1 cm, 3 cm, 5 cm에서 출아율이 10%, 8%, 5.7%로 적미에 비해 현저히 낮았으며 파종심도 7 cm에서는 출아율이 1.2%로 출아가 거의 나타나지 않았다. 이를 통해 겨울 동안의 발아력 유지에 현미 종피색이 영향을 미치는 것으로 사료되지만 본 시험에서는 매몰된 종자를 따로 채취하여 종자의 생존여부를 조사하지 않아 현미색이 겨울 동안 토양 속에서 종자의 유지에 영향을 미쳤는지 단순히 활력에만 영향을 미쳤는지에 대한 판단은 어려운 것으로 사료된다. 유효적산온도에 따른 파종심도 별 출아증가 정도를 분석한 결과(Fig. 5B), 파종심도 1 cm에서는 출아가 시작된 뒤 출아율 증가는 지속적으로 감소하였으며, 파종심도 3 cm 및 5 cm에서는 유효적산온도 70°C 및 84°C까지 출아율 증가 폭이 급격히 증가하여 왕성한 출아가 나타나기 위한 10°C 이상 최대 적산온도는 84°C인 것으로 사료된다.
Fig. 5.
Emergence rates of high dormancy weedy rice under the natural condition. (A) Emergence rates (%) of weedy rice at middle of May and (B) increase of emergence rate (%) according to effective accumulated temperature under different soil depth conditions. Effective accumulated temperature was calculated by sum the mean temperature above 10°C. Same letters are not significantly different at p<0.5 (Duncan’s multiple test).
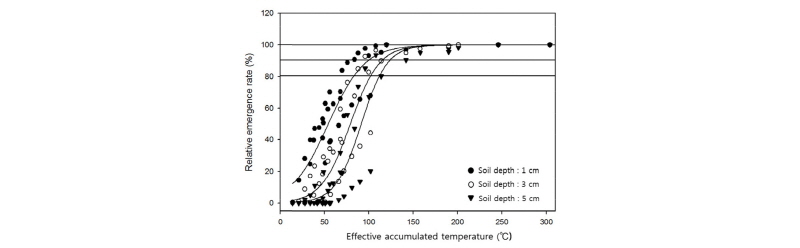
매몰깊이별 유효적산온도에 따른 상대출아율 변화분석
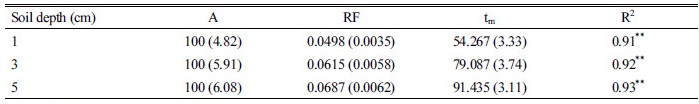
위의 데이터를 종합하여, 유효적산온도에 따른 상대 출아율 변화를 분석하였다(Fig. 6, Table 2). 최대 출아속도(RF)는 토양매몰 깊이에 따라 감소하는 경향으로 매몰 깊이 1, 3, 5 cm에서 각 0.0498, 0.0615, 0.0687인 것으로 나타났다. 상대 출아율이 50%에 달하는 적산온도는 토양 매몰 깊이 1, 3, 5 cm에서 각각 54, 79, 91°C로 나타나 매몰 깊이가 깊어질수록 출아속도가 감소하여 출아확보를 위해 많은 적산온도가 요구되는 것으로 나타났다. 토양 매몰 깊이에 따른 차이는 있지만 상대 출아율이 80%에 도달하기 위한 유효적산온도는 83-112°C, 상대 출아율이 90%에 도달하기 위한 유효적산온도는 100-123°C, 상대 출아율이 100%에 도달하기 위해서는 148-160°C의 유효적산온도가 요구되는 것으로 나타났다.
Fig. 6.
Changes in relative emergence rate (%) according to effective accumulated temperature under different soil depth condition. The relative germination rate was calculated from the germination rate at that point, assuming that the final germination rate was 100%. The curves in the graph were fitted to a logistic equation as shown in Table 2. Horizon lines indicate the 80% and 90% of relative emergence rate.
기존에 보고된 잡초성벼의 효율적인 방제법 중 잡초성벼 출아 후 비선택성 제초제 처리 및 로터리 작업을 통한 방제법이 알려져 있다(Kim et al., 2002; Im et al., 2004). 이를 위해서는 직파파종 전 잡초성벼의 출아율이 높은 시기를 판단 후 방제시기를 적절하게 결정하는 것이 중요하다. 위의 데이터를 바탕으로 시험이 진행된 전주지역의 잡초성벼 상대 출아율이 80% 및 90%가 되는 시기를 분석한 결과, 전주지역에서 일 평균온도가 10°C 이상으로 나타나는 시점은 4월 초순으로 유효적산온도가 80-110°C에 도달하는 시점은 4월 30일에서 5월 5일, 유효적산온도가 90-130°C에 도달하는 시점은 5월 1일에서 5월 8일로 나타났다.

