서 론
국내에서는1998년에 서산간척지에서 처음으로 sulfonylurea (SU)계 제초제 저항성 물옥잠(Monochoria korsakowii Regel & Maack)이 보고된 이후 현재까지 제초제 저항성 논 잡초의 초종 및 발생면적이 꾸준히 증가하고 있다(Lee et al., 2017). 최근에는 경남 김해지역의 벼 이앙재배
논에서 SU계 제초제에 대한 저항성 벗풀(Sagittaria trifolia L.)이 보고되었으며(Park et al., 2013) 이는 ALS 유전자 염기서열의 점돌연변이에 의해 197번째 아미노산 proline이 leucine이나 serine으로 치환된 결과임이 알려졌다(Won et al., 2015, 2017).
보풀속(Genus Sagittaria L.)은 택사과에 속하는 일년생 또는 다년생 수생식물로 전 세계에 약 100여 종이 주로 온대와 열대에 집중적으로, 그러나 거의 전 세계에 분포한다(Qingfeng et al., 2006). 국내 보풀속에는 올미(S. pygmaea Miq.), 벗풀, 보풀 3종이 자연상에서 발견되는 것으로 알려져 있다(Kim and Choi, 2007). 이들 3종 중 올미는 잎 중앙부가 갈라지는 화살엽을 갖지 않는 뚜렷한 특징에 의해 쉽게 구분된다. 그러나 벗풀과 보풀은 잎의 특징에서 개체변이가 매우 심하게 나타나고 그 변이가 연속적이라 두 종을 잘못 동정할 가능성이 매우 높다. Shin et al. (1997)은 두 종을 옆폭, 상부 열편 엽선의 각, 하부 열편 엽저의 형태와 같은 잎의 형태적 형질을 수리분류학적으로 조사한 결과 이들 형질은 두 종을 동정하는데 있어 유의성이 있으나 변이폭도 겹치는 것으로 조사되어 잎 형질만으로 두 종을 동정하면 잘못 동정할 가능성이 높음을 시사하였다. Shiga et al. (2017)은 실제 국내 및 일본의 표본관에서 발견된 보풀 표본에서 오동정된 표본이 많았다고 하였다. 또한, 국내외 여러 문헌을 검색한 결과 두 종의 가장 큰 비교형질은 괴경을 갖는 포복경과 엽액의 구경의 유무로 조사되었다(Kim and Choi, 2007; Qingfeng et al., 2006; Ohwi, 1984). 그러나 이와 같은 형질은 정확한 형질의 차이를 이해하지 못하는 상태에서 시료를 채취할 경우가 많아 여전히 잘못 동정할 가능성이 있다.
국내 농경지 문제잡초의 바코드 DB를 구축하기 위하여 벗풀을 채집하던 중 발견된 벗풀의 형태적 변이 폭이 매우 넓어 두 종의 정확한 동정이 필요하다고 판단되었으며 또한, 기존에 발표된 벗풀이 보풀(S. aginashi Makino)일 가능성도 배제할 수 없었다. 따라서 국내 농경지에서 채집된 Sagittaria속 잡초의 Internal transcribed spacer (ITS) 염기서열을 비교하여 두 종을 동정하고자 본 연구를 실시하였다.
재료 및 방법
본 연구에 이용된 식물재료는 국내 경기 여주, 포천, 양평, 전북 장수, 충북 증평, 괴산, 경남 하동 논에서 2017년 6-9월에 채집하여 DNA를 분리하고 확증표본을 제작하였다(Table 1). 채집된 재료의 잎의 변이 양상을 파악하기 위하여 33장의 잎을 대상으로 Shin et al. (1997)에 의해 각 분류군간 유의성 있는 차이가 있는 형질인 하부 열편의 폭, 잎의 양쪽 오목한 두 곳 사이의 거리와 상부 열편 엽선의 각을 측정하였다.
바코드 분석을 위하여 DNA는 DNeasy Plant Mini Kit (Qiagen, USA)를 이용하여 제공되는 방법에 따라 분리하였다. ITS PCR 증폭반응은 20 ng의 genomic DNA, 0.2 mM의 dNTP, 50 mM의 KCl을 포함하는 1X PCR Buffer, 10 pmole의 ITS5, ITS4 primer (White et al., 1990) 그리고 1 unit의 Taq DNA polymerase를 혼합하여 수행하였다. 증폭반응은 T-100 thermal cycler (BIO-RAD)를 이용하여 95°C에서 5분간 pre-denaturation 후, 95°C에서 1분간 denaturation, 50°C에서 1분간 annealing 그리고 72°C에서 1분간 extension을 40회 수행하였다. 염기서열분석은 GenoTech (Daejeon, Korea)에서 수행하였다.
분석된 염기서열은 Lasergen 8.1.5 (CodonCode Corp, Dedham, MA, USA)를 이용하여 contig를 작성하고 BioEdit 7.1.3의 Clustal W를 이용하여 정렬한 후 육안 편집을 거쳤다(Hall, 1999). 본 연구에서 분석된 염기서열을 비교하기 위하여 미국 국립생물공학정보센터(National Center for Biotechnology Information, NCBI)에서 보풀과 올미의 ITS 염기서열을 내려 받았으며 accession 번호는 보풀 LC278398, LC278400, LC278401, LC169553, 올미 JF978484, 벗풀 JF978494 이었다. 내려 받은 염기서열은 MEGA 6.06 (Tamura et al., 2013) 프로그램에서 Clustal W를 이용하여 본 시험에서 분석된 염기서열과 함께 정렬하였다. Kimura 2-Parameter (K2P) distance model옵션으로 종간 변이를 산출하고 Neighbor-joining dendrogram을 작성하여 벗풀과 보풀의 유연관계를 확인하였다.
본 연구에 이용된 벗풀의 확증표본은 국립농업과학원 표본관(HCCN)에 보관하였으며 ITS 바코드 염기서열은 NCBI에 제출하여 GenBank 번호를 부여 받았다.
결과 및 고찰
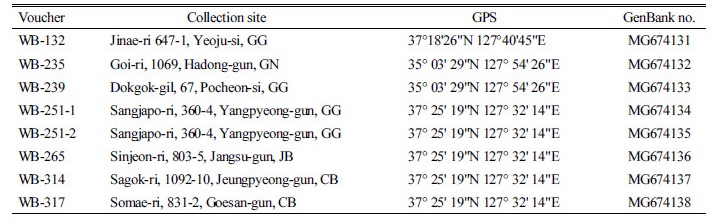
Shiga et al. (2017)이 조사한 KH 표본 중 잎의 폭이 좁은 많은 벗풀이 보풀로 잘못 동정되어 있었듯이 국내 농경지에서 채집한 벗풀의 엽형은 잎의 폭이 좁은 것에서부터 넓은 것까지 매우 심한 형태적 변이를 보였다(Fig. 1, Table 2). Shin et al. (1997)은 잎의 하부 열편의 폭 등 7개 형질을 측정하고, 이들 형질들 간의 비율 형질 등 총 11개의 형질을 이용하여 주성분 분석을 한 결과 잎의 오목한 두 지점 사이의 거리, 하부 열편의 폭, 상부 열편의 엽선 각이 높은 양의 값을 갖는 형질이라고 하였다. 이와 같은 세 가지 형질을 조사한 결과 잎의 오목한 두 지점 사이의 거리는 0.7-4.5 cm, 하부 열편의 폭은 0.3-2.0 cm, 상부 열편의 엽선 각은 20-65° 이었다(Table 2). 그러나 이들 세 가지 형질 모두 변이 폭이 뚜렷이 구분되지 않아 보풀속 식물을 분류하는 중요한 형질로 이용할 수 없음을 확인하였다(Fig. 2).

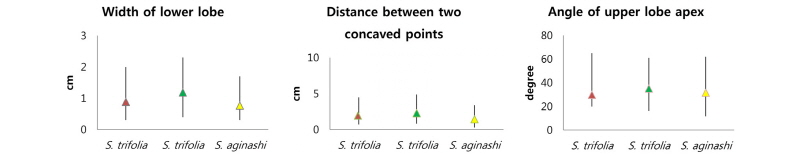
Fig. 2. Range and mean for three morphological characters of Sagittaria trifolia and S. aginashi in Korea. The lengths of bars indicate the range of leaf variation observed. Red triangle is the mean of S. trifolia measured in this study. Green and yellow triangles indicate the mean of each character of the species obtained from Shin et al. (1997).
Table 2. Ranges and means for three characters of S. trifolia and S. aginash used in this study and Shin et al. (1997).
|
|
|
yThis study. zShin et al. (1997). |
|
이와 같이 개체간에 형태적 변이가 매우 심하여 종내 또는 종간 분류군간의 한계가 모호하고 정확한 동정이 어려운 경우, 형태에 의존하지 않는 바코드 기법을 이용하면 비교적 쉽게 종을 동정 할 수 있다. 따라서 벗풀의 정확한 동정을 위하여 바코드 기법을 이용하였다. 본 종의 바코드 표준 마커인 rbcL과 matK는 NCBI에 등록되어있는 정보가 없어서 핵 DNA의 5.8S ribosomal RNA를 포함하는 ITS 부위의 염기서열을 이용하였으며 분석한 염기서열은 NCBI에 제출하여 GenBank (MG674131-MG674138) 번호를 받았다.
분석에 이용된 국내수집 벗풀 8점의 5.8S ribosomal RNA를 포함하는 ITS 염기서열은 671 bp 이었으며 개체간의 종내 염기서열 변이는 없었다. 이는 NCBI에서 내려 받은 벗풀(JF978494)의 염기서열과도 100% 일치하여 전체 채집종은 형태적 변이와 상관없이 모두 종내 변이가 없는 벗풀인 것으로 확인되었다. 보풀과의 유연관계를 확인하기 위하여 NCBI에서 내려 받은 보풀의 5.8S ribosomal RNA를 포함하는 ITS 염기서열도 671 bp 이었으며 보풀 개체간 염기서열 변이는 없었다(Shiga et al., 2017). 벗풀과 보풀은 분석된 염기서열 671 bp 중에 33 bp의 염기서열 치환이 있었고, 벗풀과 올미는 28 bp, 보풀과 올미는 단지 8 bp의 염기서열 치환이 발견되었다. 종간 변이는 벗풀과 보풀간에 4.6%, 벗풀과 올미는 4.3%, 보풀과 올미는 0.12%로 벗풀과 보풀은 다른 종과의 변이보다 높은 변이율을 보임으로써 벗풀과 보풀은 유전적으로 많은 차이가 있음을 알 수 있었다.
Neighbor-joining 계통수(Fig. 3)의 유연관계에서도 분석에 이용된 모든 벗풀은 NCBI에서 내려 받은 벗풀(JF97849)과 완벽하게 하나의 clade에 계통을 형성하며, 화살엽의 유무에 의해 쉽게 구별했던 보풀과 올미가 가까운 유연관계를 나타낸 후 벗풀과 다시 유연관계를 나타내는 것으로 확인되었다. 또한, 형태적으로 화살엽을 갖는 벗풀과 보풀은 화살엽을 갖지 않는 특징으로 분류되는 올미와의 유연관계보다 더 먼 것으로 나타나는 매우 흥미로운 결과를 얻었다.
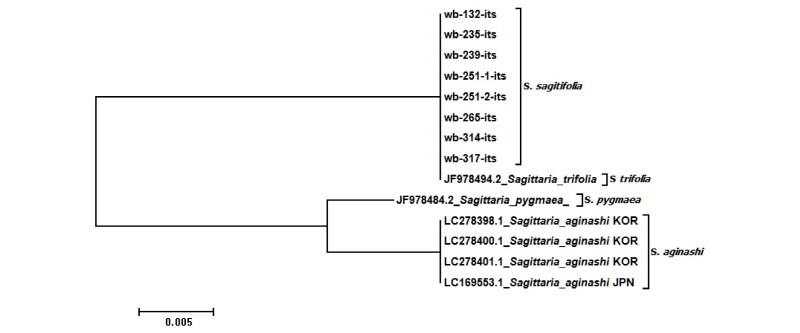
Fig. 3. Neighbor-joining tree inferred from the analysis of nrDNA ITS from Sagittaria. Branch lengths are proportional to distances estimated from the two-parameter method of KIMURA. Sagittaria trifolia showed the furtherest pairwise divergence while S. aginashi and S. pygmaea showed the closest relationship.
효율적인 잡초 관리를 위해서는, 잡초를 잘못 동정하는 경우 정확한 방제법을 놓칠 수 있으므로 정확한 종 동정이 선행되어야 할 것이다(Mennan and Kaya-Altop, 2012). 캐나다 온타리오 남서부지방에 자생하는 비름속(Amaranthus L.)은 atrazine저항성과 감수성 종이 혼재하여 종에 따라 다른 방제법이 요구되었다(Warwick and Weaver, 1980). 이와 같이, 같은 속의 잡초라 할지라도 종에 따라 다른 관리법이 필요한 경우가 있어 정확한 종 동정은 매우 중요하다. 국내에서도 제초제저항성 잡초는 꾸준히 확산되고 있으며 이들 제초제저항성 잡초의 효율적인 관리를 위해서는 정확한 종 동정이 필요하다.
진화속도가 빠른 ITS 구간은 multicopy로 존재하여 종종 클로닝을 해야 하는 어려움이 있어 고등식물의 표준 바코드마커에서 제외되었으나 클로닝없이 염기서열 분석이 가능할 경우 유용한 추가 바코드가 될 수 있다고 하였다(CBOL Plant Working Group, 2009). 보풀속과 같이, 분류군에 따라서는 ITS 만으로도 충분히 바코드가 가능한 경우가 있다. 본 연구에서도 비록 ITS가 식물의 표준 바코드 마커는 아니지만 ITS 염기서열만으로도 보풀과 형태적으로 구별하기 쉽지 않은 벗풀을 쉽고 빠르고 간단하게 동정해 낼 수 있었다. 그러므로 벗풀과 같이 형태적으로 뚜렷한 종 동정이 어려운 경우 바코드는 강력한 분류 동정대안이 될 수 있다.

