서 론
최근 급격한 도시화, 산업화에 따른 석유 자원의 고갈과 지구온난화 현상으로 인해 전 세계 각국은 대체 에너지원 개발에 매진하고 있다. 우리나라도 2015년 12월에 체결된 파리협정에서 이산화탄소를 2030년까지 37% 감축하겠다고 천명하고 신재생에너지 분야에 투자를 늘리고 있는 실정이다. 신재생에너지 중 바이오연료는 자원이 빈약한 우리나라에서도 생산이 가능하며 석유를 대체할 수 있을 정도로 유용성이 높은 재생에너지이다. 그러나 식량자원의 바이오에탄올 원료로의 이용은 국제 곡물가의 폭등, 여러 후진국에서 식량 파동과 같은 심각한 윤리적 문제를 야기 시키고 있어서 이러한 문제점을 해소하기 위하여 식량작물이 아닌 비식량작물(억새, 스위치그래스, 포플러 등)을 이용한 바이오에너지 원료 확보에 대한 관심이 증가되고 있다(Atkinson, 2009; Greef et al., 1997; Lewandowski et al., 2000; Yan et al., 2012).
그 중 억새(Miscanthus)는 화본과(Poacea), 기장아과(Panicoideae), 쇠풀족(Andropogoneae), 억새속(Miscanthus)에 속하는 식물이다(Chen and Stephen, 2006). 억새속 식물에는 17종이 존재한다고 알려져 있으며(Greef et al., 1997; Park et al., 2009), 한국, 일본, 중국 등 동아시아 전역에 널리 자생하고 있는 잡초로, 과거 가축의 조사료나 경사지의 토양 유실 방지 등에 이용되어왔다(Yu et al., 2016). 이후 비식량작물을 이용한 바이오에너지 원료로 각광받기 시작하였으며, 이 중 바이오에너지용으로 주목받고 있는 종은 한국 등 동아시아가 원산인 참억새(M. sinensis)와 물억새(M. sacchariflorus) 그리고 물억새 4배체와 참억새 2배체의 중간 교잡종인 이질 3배체 억새(M.× giganteous)등이다(Lewandowski et al., 2000; Moon et al., 2010). 현재까지 유럽, 미국 등지에서 바이오에너지용으로 가장 널리 연구되고 있는 억새는 3배체 억새이다(Yan et al., 2012; Yook et al., 2014).
또한 억새는 C4 광합성 경로를 가지는 다년생 식물이며 질소 양분이용 효율이 우수하여 무비재배가 가능하고, 일년생 작물 재배 시 요구되는 농작업 등을 생략할 수 있어서 에너지 산출 및 투입수지가 높다(Lewandowski and Schmidit, 2006; Moon et al., 2010). 억새 종과 관련하여 세계적으로 많은 연구가 이루어져 왔으며 억새의 원산지인 일본과 중국에서 매우 활발하다. 일본의 Hokkaido 대학에서는 억새 품종 개발 연구를 진행 중이며, 중국에서는 현지의 억새 유전자원 수집 및 품종개발 연구를 진행하고 있다.
우리나라에서도 억새 수집 및 형태적 특성에 관한 연구가 이루어졌는데 Yoo et al. (2012)이 국내 8종, 미국 3종, 인도, 네팔 각 1종 등 총 14종의 국내외 억새를 대상으로 형태적 특성을 조사하여 보고한 바 있다. 억새를 이용한 바이오에너지 산업의 발전과 확장을 위해서는 새로운 품종 개발이 요구되며, 그 방안으로 다양한 억새 유전자원의 확보는 억새의 유전적 기반을 넓히는 의미에서 중요하다고 할 수 있다(Jensen et al., 2013; Arnoult et al., 2014).
따라서 본 연구는 국내 각지에서 수집한 거대억새를 포함한 물억새, 참억새 등 약 960점의 억새 유전자원을 수집하고 그 형태적 특성을 평가함으로써 바이오에너지 원료작물로서의 억새 품종육성을 위한 기초자료를 축적하기 위하여 수행하였다. 특히 수집된 유전자원을 2013년에서 2015년까지 3년간 억새 유전자원 보존 시험포와 시험포장에서 재배하고 동일 조건하에서 나타나는 유전자원별 형태적 특성과 생육을 비교 검토하였다.
재료 및 방법
억새 유전자원 수집
억새 유전자원은 2010년부터 2013년까지 국내 여러 지역에서 960점을 수집하였으며 수집된 유전자원의 지역별 점수는 Table 1과 같다. Chen and Stephen (2006)이 제안한 포영, 줄기 및 지하경 특성을 토대로 물억새와 참억새로 구분하여 수집하였고 물억새는 551점, 참억새는 409점이었다. 물억새는 불규칙한 형태로 줄기가 형성되며 늦가을 및 겨울에는 줄기의 잎집이 탈락되고, 지하경은 길고 가늘며 넓게 뻗어나가며 소수(spikelets)에는 솜털(callus hair)이 있으나 까락(awn)은 없는 것이 특징이다. 참억새는 한 개체가 조밀하게 군락을 이루어 줄기가 형성되며 늦가을 및 겨울에도 잎집이 탈락되지 않고, 지하경은 짧고 서로 촘촘하게 엉켜있으며, 소수에는 솜털 및 까락이 존재한다(Moon et al., 2010; An et al., 2015). 이러한 특성에 기초하여 억새 한 줄기에서 하나의 지하경을 굴취하였고 비닐 지퍼백에 밀봉하여 운반하였다. 수집할 때 지역의 위도, 경도, 고도를 조사하였으며 형태적 특성으로는 초장과 경태, 마디수를 측정하였다.
억새 유전자원의 재배
각 지역에서 수집된 억새 유전자원의 지하경은 전남 무안군에 위치하는 국립식량과학원 바이오에너지작물연구소로 운반 후 억새 한 개체씩 PVC 통(L: 120 cm, × W: 72 cm × H: 35 cm)에 재식하였다. 재식 후 각 유전자원의 특성이 나타날 수 있도록 비료 시용은 하지 않았으며 재배 기간 동안 매년 2월 중 ․ 하순경에 지상부를 모두 제거하였다. 기타 재배관리는 모든 유전자원에 대하여 동일한 조건이 적용될 수 있도록 관리하였다. 억새 유전자원 보존포장 내의 억새 생육 상황은 Fig. 1과 같다.

억새 유전자원의 형태 및 생육 특성 평가
수집된 억새 유전자원을 동일한 생육조건에서 재배시키면서 형태 및 생육 특성을 평가하였다. 수집된 960점의 유전자원에 대한 수집 전후의 생육변이를 검토하기 위하여 수집 현지에서의 생육 특성 조사와 연구소 재배포장에서의 생육특성 조사가 모두 이루어진 유전자원 463점을 선정하여 재배 3년차 초장 성적으로 비교 분석하였다.
형태 및 생육 특성의 정밀한 조사를 위하여 960점의 유전자원 중 61점의 대표자원을 다음의 기준으로 선발하였고 이후 특성평가에 사용하였다. 특히 생산량이 우수하여 신품종 육성 시 활용할 수 있거나, 물억새임에도 참억새의 특징을 갖는 유전자원을 선발하였다. 물억새 유전자원의 선발기준은 건물중 1 kg 이상의 생육이 우수하거나 생육이 불량한 것과 동계시기에도 잎집이 붙어 있는 것, 참억새와 같이 줄기가 촘촘히 자라는 형태의 것(참억새 형질), 잎이 가늘고 뾰족한 형태를 나타내는 것, 조기 출수하는 것을 선발하였다. 참억새 유전자원은 건물중이 2 kg 이상의 것, 조기 출수하는 것, 도복성이 강한 것 등을 기준으로 선발하였다. 경관용 억새는 조기 출수하는 유전자원이 대부분이었는데 그 중에서 잎에 줄무늬가 있거나 잎이 뾰족하고 곧게 자라는 것을 선발하였다, 또한 국내 유전자원에 비하여 줄기가 통통하고 생육량이 월등이 많은 중국에서 수집한 억새 유전자원을 비교군으로 포함시켰다. 선발 자원수는 물억새(M. sacchariflorus) 29점, 참억새(M. sinensis) 23점, 경관용억새 4점, 이질 3배체 억새(M. × giganteous) 1점, 중국 수집종(M. lutarioriparius) 4종이었다.
선발된 유전자원은 3월말부터 4월초까지 맹아일, 6월말부터 11월초까지 지엽전개일 및 출수일을 조사하였으며, 생육이 활발한 시기에는 잎길이, 잎너비, 잎색, 잎 직립성, 엽이색, 엽이유무 등을 조사하였다. 억새의 생육말기인 11월경에 초장, 경태, 생육일수, 건물중량, 이삭길이, 까락유무, 줄기노화도, 도복유무, 줄기마디색 등을 조사하였다.
선발된 억새 유전자원의 수집지역 정보는 Table 4와 같다. 물억새 유전자원은 총 29점으로 전남에서는 거대억새류를 포함한 6점, 전북에서는 2점, 경남 4점, 경북 4점, 충남 9점, 충북 2점, 경기 2점이었다. 참억새 유전자원은 총 23점이며 제주도에서 수집한 유전자원이 가장 많은 15점이 선발되었다. 경관용억새는 참억새의 일종으로 알려져 있으며, 총 4점(Morning light, Little zebra, Yakushima dwaft, Little kitten)이었으며, 중국의 양쯔강에서 수집한 유전자원은 억새종 분류 중에서 M. lutarioriparius로 알려져 있으며 이 중 4점을 선발하였다.
유연관계 분석
선발된 유전자원의 형태적 특성 분석은 NTSYS-pc program Version 2.1을 사용하여 수행되었다. 유전자원의 형태적 특성 중 잎색, 직립성, 엽이, 엽이유무, 까락유무, 줄기노화도, 도복유무, 줄기마디색을 조사하여, 본 조사 항목들이 관찰되면 ‘1’, 관찰되지 않으면 ‘0’으로 표시하여 매트릭스를 작성하였다. 나머지 형태적 특성인 잎길이, 잎너비, 초장, 경태, 건물중량, 이삭길이, 출수율은 선발된 전체 유전자원의 평균 이상이면 ‘1’, 평균 이하에서는 ‘0’으로 표시하였다. 이러한 매트릭스의 데이터 분석은 표준화(standardization) 작업 후, Jaccard’s similiarity coefficient (Jaccard, 1912)에 기초하여, 각 유전자원들의 유의성(similarity)를 분석하였다. 또한 분석된 결과를 기초로 clustering 분석을 통해 phylogenic tree를 작성하여 유전자원간의 유연관계를 분석하였다.
결과 및 고찰
억새 유전자원 재배 기간 중의 기온 및 강수량
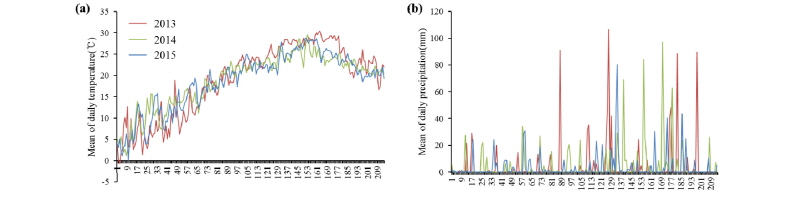
2013년부터 2015년까지 3년간 억새의 맹아가 시작되는 3월 1일부터 억새의 생육말기인 9월 30일까지 총 214일 동안의 재배지인 무안지역의 일평균 기온과 일평균 강수량은 Fig. 2와 같다. 2013년의 평균기온은 19.0°C였으며, 최고기온은 8월 22일에 45.5°C였고, 2014년의 평균기온은 18.7°C였으며, 최고 평균기온은 7월 31일에 35.5°C, 2015년의 평균기온은 18.7°C였으며, 최고 평균기온은 8월 7일에 35.6°C였다. 평균기온과 최고 평균기온은 2013년이 가장 높았던 것으로 나타났다. 작물의 생육에 중요한 수분 공급량을 파악하기 위해 연도별 일평균 강수량과 연간 누적 강수량을 확인하였다. 2013년의 일평균 강수량은 4.2 mm이었으며 누적강수량은 893.5 mm였고, 2014년의 일평균 강수량은 4.7 mm, 누적 강수량은 999.5 mm이었고, 2015년의 일평균 강수량은 3.2 mm, 누적강수량은 685.5 mm이었다. 일평균 강수량 및 누적 강수량은 2015년이 가장 적었으나, 3년간 강우일수는 2013년에는 51일, 2014년에는 69일, 2015년에는 67일로 나타나 2013년의 강우일수가 가장 적었다.
억새 유전자원의 형태적 및 생육특성 분석
수집된 960점의 억새 유전자원 중 수집 현지에서 특성조사가 이루어진 자원 463점에 대하여 수집 현지에서의 초장과 연구소 포장에서 3년간 재배한 후의 초장을 각각 측정하여 비교 분석하였다(Table 2).

유전자원을 수집할 때 달관으로 선발된 개체의 초장을 측정하고 지하경을 굴취하여 연구소 포장에 재식하였다. 재배 3년 후 각각의 초장을 측정하여 3년 전의 수집 현지 초장 성적과 동일 조건에서 재배한 후의 초장 성적의 변화를 분석하였다. 일반적으로 억새를 3년 재배하면 적정 생산시점에 도달하는 것으로 보고되었다(Moon et al., 2010). 따라서 3년 재배 후 초장이 수집현지 초장보다 커진 경우는 Higher (+), 작아진 경우는 Smaller (-)로 표현하였다. 조사된 463점의 유전자원 중 38.4%인 178종은 초장이 더 커졌으나 285종(61.6%)은 오히려 초장이 줄어들었다. 이는 토양, 기후 등 환경에 따른 반응 차이로 이런 결과가 나타난 것으로 생각된다. 일반적으로 자생지는 식물의 특성이 제대로 발휘되고 있는 지역으로 여겨져 유전자원 수집시에는 당시의 생육 상태를 보고 유전자원을 수집하게 된다. 그러나 본 연구 결과, 우수자원 선발 및 대단위 재배를 위한 목적으로 현지의 생육 상태만으로 선발하는 것을 위험할 수 있다는 것을 알 수 있었다.
물억새의 초장 생육 감소비율은 75.2%였으며, 참억새는 오히려 수집지역에서의 초장보다 더 커진 비율이 58.2%로 많았다. 이 결과는 참억새와 물억새 고유의 특성에서 기인한 바가 큰 것으로 사료된다. 참억새는 주로 마른 땅에 자생하지만 물억새는 강 또는 호수 주변을 따라 분포하고 있다(Christian and Haase, 2001; Kim et al., 2004). 이러한 물억새를 수변 자생지에서 평야지로 옮겨 심어 토성, 토양 수분 등 토양 환경이 변화하게 되어 초장이 감소한 것으로 사료된다.
참억새의 수집지역별 생육변화 또한 이러한 결과를 뒷받침 해준다. 참억새는 189종의 수집종 중 제주도에서 146종, 기타 지역에서 43종이 수집되었는데 이식 후 초장이 늘어난 수집종은 제주 수집종에서 65.8%였던 반면 기타 지역 수집종에서는 32.6%에 불과하였다(Table 3).

일반적으로 참억새는 배수가 잘되는 산록경사지에서 자생한다. 척박지에 적응성이 높아, 바위절벽 틈과 같은 매우 열악한 환경에서도 생육할 수 있으나, 습해를 입을 수 있는 토양 조건에서는 잘 자라지 못한다(Christian and Haasse, 2001; Moon et al., 2010). 제주도 자생 참억새는 습해를 입을 수 있는 수분이 많은 토양에서 자생하고 있었으나, 연구소로 이식 재배하였을 시 토양 수분이 줄어든 평야지에서 재배되었다. 이는 습해를 입을 수 있는 조건에서 참억새의 고유 특성에 맞는 생육조건에 이식 재배됨으로써 초장이 커졌을 것으로 사료된다.
식물은 종류에 따라 생육에 적합한 환경조건에 차이가 있으며 자생식물은 이러한 생육환경 조건이 알맞은 곳에서 군락을 이루게 된다(Lim et al., 1990; Eom and Kim, 2017). 따라서 자생식물의 재배화를 위해서는 자생지의 환경과 생육 상태를 고려하여 최적 조건을 맞추어 주어야 하는데, 수집된 유전자원을 환경 조건이 고정된 재배지에서 재배할 경우에는 자생지의 생육특성이 나타나지 않을 수 있다는 것을 고려해야 할 것이다.
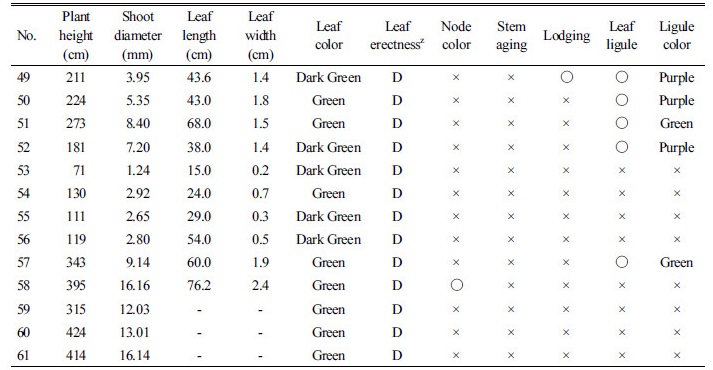
수집된 960점의 억새 유전자원 중 선발된 억새 유전자원 61점의 수집 지역 및 형태적 특성을 조사하였다(Table 4, 5). 물억새의 초장은 183-319 cm였으며, 전남지역에서 수집한 억새의 초장과 경태가 가장 크거나 두꺼운 것으로 나타났다. 반면 경북 안동에서 수집한 유전자원의 초장과 경태는 가장 작거나 얇게 나타났다. 물억새 유전자원의 잎색은 대부분 녹색이었으나, No. 17과 19는 진녹색이었다. 잎집 탈락여부를 조사한 결과 물억새인 No. 17, 22, 24, 26, 28, 29는 생육말기에도 잎집이 떨어지지 않고 줄기를 싸고 있었다. 또한 까락의 유무를 조사한 결과, 대부분의 물억새 유전자원들은 까락이 없었으나 경북 문경에서 수집된 No. 19은 까락이 있는 것으로 조사되었다. 물억새 No. 16은 조사된 유전자원 중 가장 일찍 출수하였는데 이는 충남 태안읍에서 수집된 것으로 맹아일은 4월 4일이었으며, 지엽전개일은 7월 13일, 출수일은 7월 30일이었다. 거대억새 1, 2호로 알려진 No. 15와 No. 13의 지엽전개일은 9월 4일, 5일이었으며 출수일은 각각 9월 16일, 18일로 나타나 가장 늦게 출수되었다.
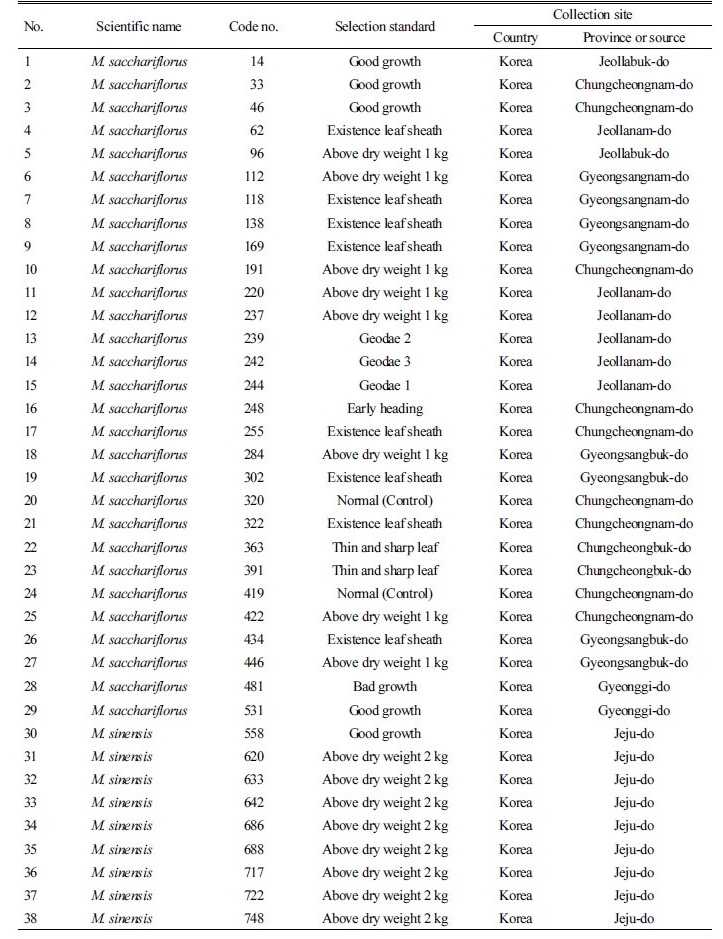
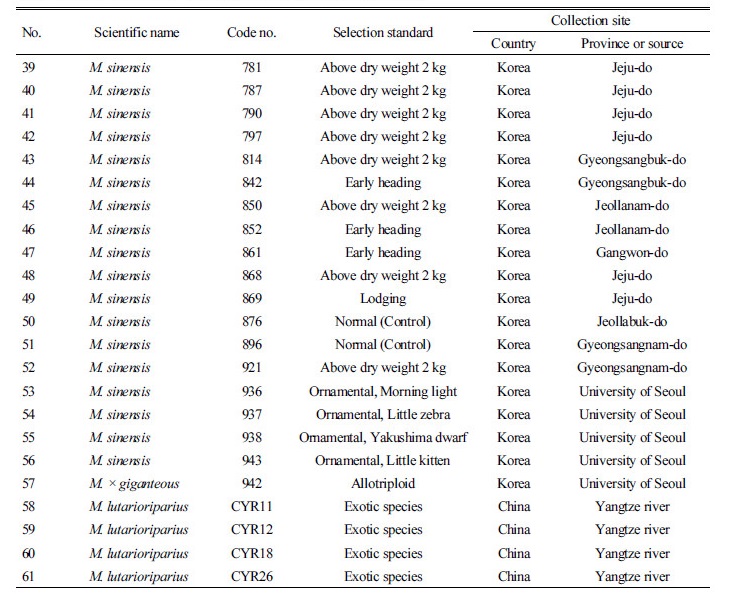
Table 5. Morphological characteristics of 61 Miscanthus germplasms (Continued).
|
|
|
zD: Direct; E: Erect type; ○: Existence; ×: Nonexistenc. |
|
참억새의 초장은 181-277 cm였으며, 경태는 3.95-8.73 mm이었고 그 중 제주도 신흥리에서 수집한 유전자원의 초장 및 경태가 가장 크거나 굵은 것으로 나타났다. 잎색은 대부분이 진녹색이었으며 대부분 억새 생육말기에도 잎집이 떨어지지 않고 줄기를 싸고 있었다. 모든 참억새 유전자원은 까락이 존재하였다. 참억새의 일종으로 알려진 경관용 억새(No. 53, 54, 55, 56)는 초장, 경태, 엽장, 엽폭 등이 다른 억새에 비하여 작았으며 지엽전개일 및 출수일은 가장 빨랐다. 경관용 억새는 참억새류로 분류되었으며 모든 경관용 억새의 이삭에는 까락이 존재하는 것으로 나타났다.
중국에서 수집된 억새종의 초장은 315-424 cm였으며 경태는 12.03-16.16 mm로 조사된 유전자원 중 초장 및 경태가 가장 크거나 굵은 것으로 나타났으며, 잎길이 및 잎너비도 가장 길었다. 생육 및 형태적 특성 조사에 의한 억새 유전자원의 특성 조사 결과, 같은 물억새 및 참억새 중에서도 형질이 매우 다양하게 분포하고 있음을 알 수 있었다.
조사된 억새의 생육특성 및 형태적 특성 분석의 결과를 바탕으로 각 개체간의 correlation coefficient를 구한 뒤, phylogenic tree를 작성하였다(Fig. 3). 61종의 유전자원은 크게 두 그룹으로 나뉘어졌으며, 물억새 그룹(Group I) 31종과 참억새 그룹(Group II) 30종으로 구분되었다. 물억새 유전자원은 크게 Group I-1과 Group I-2로 나뉘었으며, Group I-1에는 No. 1 등 27종이 Group I-2에는 No. 6 등 4종이 포함되었다. 물억새 Group I-1은 Group I-1a와 Group I-1b로 최종 나누어졌으며, Group I-1a에는 No. 1 등 18점이, Group I-1b에는 No. 5 등 9점이 포함되었다.
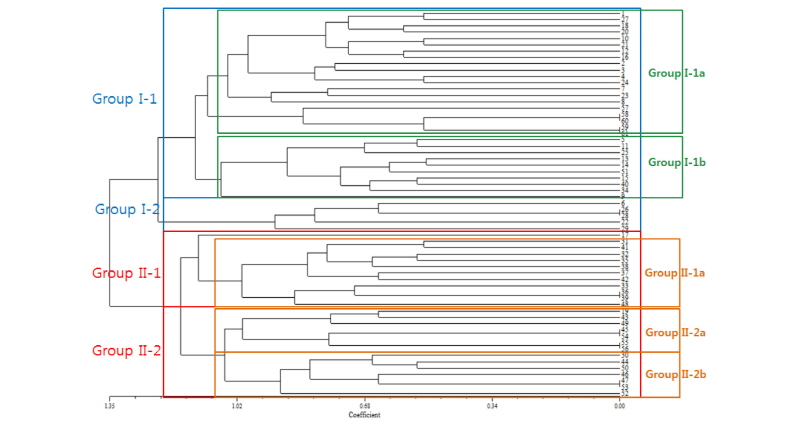
Fig. 3. Phylogenic tree of 61 Miscanthus germplasms. It is divided into two classes: Miscanthus sacchariflous (Group I, Sky line) and Miscanthus sinensis (Group II, Red line). Green line and orange line represent the classification within group I and the classification within group II, respectively. Phylogenic tree was created using NTSYS-pc program Version 2.1.
흥미롭게도 No. 57, 58, 59, 60, 61 등 유전자원들은 이질 3배체 억새(No. 57)와 중국 수집종(No. 58, 59, 60, 61)이었는데 Group I-1a로 분류되었다. Group I-1b에는 No. 5 등 9점의 유전자원이 포함되었으며, 이 중 No. 13, 14, 15는 국내 1호 억새 품종인 거대억새 1호(No. 15)의 수집지역에서 수집된 자원이었다. 물억새 Group I-2는 No. 6 등 4종의 자원이 포함되었으며 참억새 유전자원(No. 34, 40)과 경관용 억새 유전자원(No. 51)이 일부 포함된 것을 알 수 있었다.
참억새의 유전자원은 Group II-1과 Group II-2로 분리되었으며, Group II-1은 11점이 포함되었으며, Group II-1의 10점과 이에 포함되지 않은 No. 17 1점으로 분리되었다. No. 17은 Group II-1에 속해 있지만 Group II-1a와의 유연관계는 먼 것으로 나타났다. 이 유전자원은 물억새 계열에 속하는 유전자원으로서 다른 물억새와 달리 잎집이 존재하였으며 잎색은 진녹색을 나타내는 등 형태적으로 참억새 형질을 띄고 있었으나, 다른 참억새들이 까락이 존재하는 반면 까락이 없는 특성을 가지고 있어 참억새와 물억새의 특징이 혼재되어 있음을 알 수 있었다.
Group II-2는 Group II-2a 5종과 Group II-2b 6종으로 분류되었다. Group II-2a 중 No. 19는 잎색은 진녹색이며 까락이 존재하여 물억새와 참억새의 특징을 동시에 갖고 있어 phylogenic tree 분석 결과 참억새 그룹으로 포함되었다. Group II-2a 중 No. 54, 55, 56은 경관용 억새였다.
본 연구 결과를 종합해본 결과, 억새 종들의 고유 특성으로 인하여 수집 유전자원 중 참억새보다 물억새의 유전자원들이 자생지에서 보다 초장이 감소하였으며, 참억새 중 주로 제주도에서 수집된 유전자원들의 초장이 증가하였다. 유전자원간 다양성을 평가한 결과, 크게 참억새와 물억새로 나뉘었으나, 참억새로 분류된 일부 자원들이 평가 후 물억새로 분류되거나 물억새로 평가되던 자원들이 참억새로 분류되는 경우도 존재하였다. 이는 물억새와 참억새의 특성이 혼재되고 있는 유전자원임을 확인하였다. 향후 이러한 유전자원을 대상으로 배수성 검정 등 종 동정을 통해 특성이 혼재되고 있다는 점을 유전적 차원에서 검증해야 할 것이다. 이러한 유전자원들은 우수한 바이오매스 작물의 육종을 위한 중요한 재료로 활용 할 수 있으며, 본 연구 결과는 우수한 자원을 육성하기 위한 육종 조합을 위한 기초 자료로 유용하게 활용할 수 있을 것으로 사료된다.

