서 론
기후변화와 국제 교역의 확대 등으로 외래 잡초 발생의 위험성이 증가하고 있다(Oh et al., 2002). 2001년까지 우리나라에 유입된 외래잡초는 37과 315종이며 주로 도로 주변 등과 같은 비농경지에서 발생되고 있는 것으로 조사되었다(Oh et al., 2002). 그러나 최근 우리나라 밭작물 재배지 잡초발생 및 분포현황(Lee et al., 2015), 제주도 밭의 외래잡초 분포(Kim et al., 2015a), 농경지 내 생태계교란식물의 분포(Kim et al., 2017) 등의 조사에서 외래잡초가 농경지에서도 발생이 증가하는 것으로 확인되었다. 생태계교란식물들은 생태계의 균형을 교란하거나 우려되는 식물로 환경부장관이 지정한다(Kil et al., 2012). 현재 생태계교란식물은 단풍잎돼지풀(Ambrosia trifida L.), 가시박(Sicyos angulatus), 가시상추(Lactuca scariola) 등을 포함한 14종이 지정되었다(Kil et al., 2012; Kim et al., 2017). 이들 생태계교란식물은 주로 종자 생산량도 많고 초기생장이 빨라 경합력이 높기 때문에 생물다양성을 훼손시킬 뿐만 아니라, 인간 활동을 제한하며 건강을 위협하는 요인이 되고 있다(Andow, 2003).
외래잡초 둥근잎유홍초(Quamoclit coccinea Moench)는 “능조라”라고도 불리며 메꽃과의 일년생 덩굴식물로 원산지는 열대아메리카이며 원예종으로 국내에 도입되어 전국 각처의 인가에서 관상초식물로 사용되기도 하였다(Kim et al., 2015b; Yang et al., 2004). 둥근잎유홍초의 형태적 특징은 초장이 2-3 m 내외로 자라며 줄기는 왼쪽으로 감고 올라가며 밑을 향한 털이 있고, 잎은 어긋나며 심장형으로 보통 3개로 갈라지고, 잎자루는 3-4 cm로 길다. 최근에는 농경지와 비농경지에 대량 발생하여 피해를 주는 외래잡초이다.
둥근잎유홍초는 현재 생태계교란식물로 지정되지는 않았지만 생육이 왕성하고 신속하며 종자 생산량도 많아 제거방법이 단순하지 않다. 또한 병해충의 서식처를 제공하고, 인체 및 가축의 피해, 국내 토종 식물생장 방해 그리고 생태계 파괴 유발 등 많은 피해와 농작물 수량감소까지 영향을 미치고 있는 실정이다(CBD, 2009). 우리나라에서 둥근잎유홍초는 콩밭과 같은 밭작물재배지에서 발생량이 증가하고 있다. 일본에서도 콩 재배지에서 둥근잎유홍초의 발생이 큰 문제시되고 있다(Seo, 2010). 따라서 본 연구는 다양한 온도, 파종심도, 차광 및 토양수분 조건이 둥근잎유홍초의 발아와 생육에 미치는 영향을 구명하여 효과적인 관리방법을 개발하는데 필요한 기초 자료로 사용하고자 수행하였다.
재료 및 방법
본 실험에 사용된 둥근잎유홍초(Quamoclit coccinea Moench) 종자는 전라남도 순천시 순천대학교 서면 농장에서 2016년 11월에 채집하여 종자를 정선하여 실험에 사용할 때까지 4°C 저온냉장고에서 보관하였다.
둥근잎유홍초의 종자 발아율
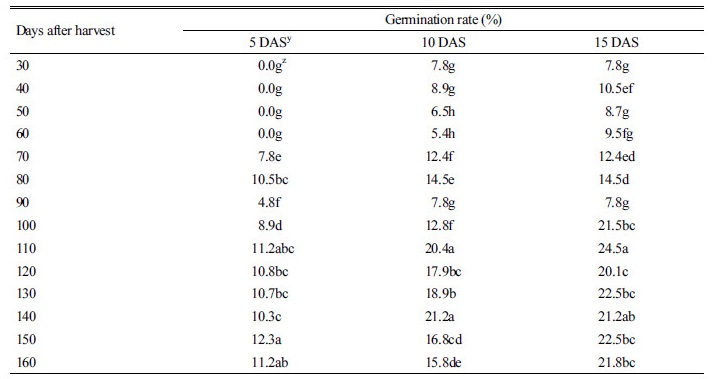
저온처리(4°C 냉장고)한 둥근잎유홍초 종자의 발아율을 확인하기 위하여 수확 후 30일부터 10일 간격으로 160일까지 조사하였다. 종자의 발아율 실험은 일회용 멸균 petri dish (90 mm)에 여과지(Whatman paper 2, Whatman, Inc., Maidstone, UK) 2장을 놓은 다음 증류수 5 ml를 첨가하고 각 petri dish 당 10립씩 치상하여 5, 10, 15일 후에 발아율을 조사하였다. 생장상의 조건은 온도 25±2°C, 상대습도 70% 그리고 광주기 12/10시간(주/야), 광량(100 µmol m-2 s-1)로 하였다. 파종 후 3일까지 암상태로 유지하고 그 후에는 위의 광주기 상태로 유지하였다.
화학약품 처리에 의한 둥근잎유홍초의 종자 휴면타파
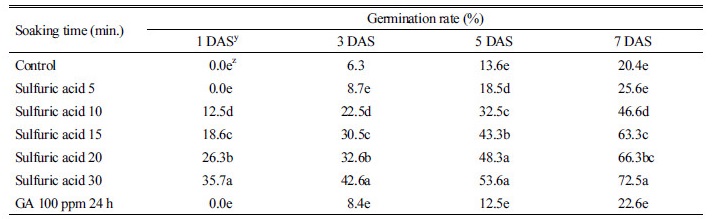
위의 종자 발아율 실험에서 둥근잎유홍초의 종자가 발아율이 저조하였으므로 농황산(97%)에 0, 5, 10, 15, 20, 30분간 침지처리 한 후 2시간 동안 흐르는 수돗물로 수세하거나 100 ppm 지베렐린에 24시간 침지 한 후 곧바로 수세하고 위의 “종자 발아율” 실험과 동일한 방법으로 발아율을 조사하였다.
온도조건별 둥근잎유홍초 발아율 및 생육조사
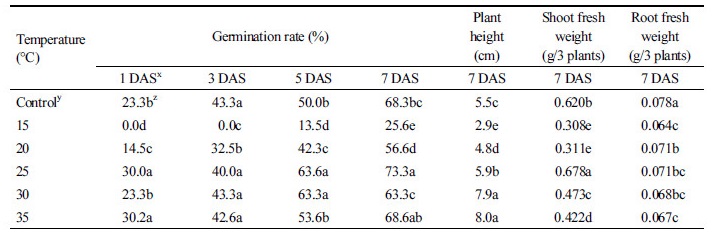
온도조건별 발아특성을 구명하기 위하여 둥근잎유홍초 종자를 농황산에 15분간 침지하여 휴면타파한 후 15, 20, 25, 30, 35°C 생장상에 치상하였다. 생장상 조건과 발아율 실험은 위의 “종자 발아율” 실험과 동일하게 하였고 온도만 다르게 설정하였다. 파종 1, 3, 5, 7일 후 발아율과 파종 7일후에 초장, 지상 및 지하부 생체중을 조사하였다.
파종심도별 둥근잎유홍초의 발아 및 생육 조사
농황산에 15분간 침지하여 휴면타파한 종자를 사양토로 충진된 폿트(지름 100 mm×높이 300 mm)에 0, 0.5, 1, 2, 3, 5, 7, 8, 10, 12, 15, 17, 20 cm 깊이로 파종한 후 위의 “종자 발아율”실험과 동일한 생장상을 이용하여 파종하고 1, 5, 10, 15일 후 발아율과 파종 15일 후에 중배축 길이, 초장 및 지상부 생체중을 조사하였다.
차광정도별 둥근잎유홍초의 발아 및 생육 조사
본 실험은 바람막이 시설이 설치된 야외에서 수행되었다. Pot (지름150 mm×높이150 mm)에 사양토를 넣은 후 농황산에 15분간 침지하여 휴면타파한 종자를 0.5 cm 깊이로 파종하였다. 파종 후 자연광 조건과 자연광의 20, 35, 50, 75, 90%로 차광되도록 차광막을 설치하였다. 실험기간 중 온도는 주/야 30±2°C/20±2°C이었고, 강우는 없었고 적절한 토양수분을 유지하였다. 파종 후 3, 5, 10, 15일에 발아율과 파종 후 15일에 엽수, 초장 및 생체중을 조사하였다.
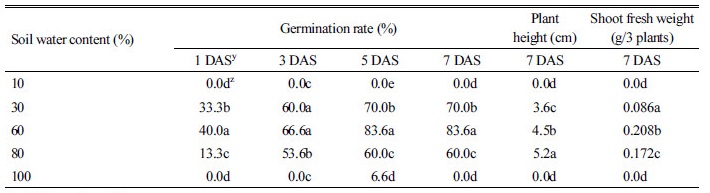
토양수분 함량별 둥근잎유홍초의 발아 및 생육 조사
본 실험에 사용된 토양(사양토)은 60°C에서 2일간 건조한 후 토양 무게를 달아 pot (지름 70 mm×높이 70 mm)에 넣은 후 토양수분을 10, 30, 60, 80, 100% (포화상태)로 조절하고 농황산에 15분간 침지하여 휴면타파한 종자를 0.5 cm 깊이로 파종하였다. 파종 후 1, 3, 5, 7일에 발아율과 파종 후 7일에 초장 및 생체중을 조사하였다. 실험기간 중 매일 줄어든 수분을 보충하여 주었다.
통계처리
본 실험은 완전임의 배치 6반복으로 하였으며, 실험결과는 SAS (2000) 통계분석 최소유의차 검정(P=0.05)을 실시하여 유의성 유무를 확인하였다.
결과 및 고찰
둥근잎유홍초의 종자 휴면
둥근잎유홍초의 종자 휴면 정도를 알아보기 위하여 수확한 종자를 4°C 저장고에 보관 후 30일부터 10일 간격으로 160일까지 발아율을 조사하였다(Table 1). 둥근잎유홍초의 종자 수확 30-90일후에 발아율은 8-15%였고, 100-160일 후에는 20-23%였으며, 발아에 소요되는 일수도 10일 이상이었다. 또한 수확 후 1년이 경과한 종자의 경우도 여전히 20-30% 이하의 발아율을 보였다(자료 미제시). 따라서 둥근잎유홍초 종자의 경우 휴면 정도는 최소 1년 이상으로 간주된다. 종자휴면은 잡초 속성 중 하나로서 잡초종자 또는 영양번식기관에서 관찰되고 있다(Mayer and Shian, 1974). 종자의 휴면 원인은 발육이 불완전 배, 생리적으로 미숙한 배, 기계적 저항성을 지닌 종피, 발아억제물질 존재 등의 요인으로 분류할 수 있다(Aldrich and Kremer, 1997). 따라서 이러한 요인들에 의해서 휴면은 잡초가 불량한 환경조건에서도 생존할 수 있는 특성뿐만 아니라, 시기를 달리해서 또 다른 장소에서 발아할 수 있는 특성이라고 할 수 있다. 즉 발아의 동시성 없이 둥근잎유홍초가 오랜 시일에 걸쳐 발생하게 되므로 새로운 잡초문제에 직면하게 될 수도 있다.
둥근잎유홍초의 종자 휴면타파
위의 종자 휴면실험에서 둥근잎유홍초 종자는 휴면성을 지니고 있는 것이 확인되었는바, 휴면을 타파하는 방법을 밝히는 것은 잡초를 합리적으로 방제하기 위하여 매우 중요하다. 잡초 종자의 휴면타파방법은 다양하지만 인위적인 방법에는 파상법(scarification), 저온 또는 변온을 부여하여 휴면을 타파하는 온도처리 방법, 그리고 각종 화학물질을 사용하여 휴면을 타파하는 약품처리 방법 등이 있다(Aldrich and Kremer, 1997). 본 연구에서는 둥근잎유홍초 종자는 저온에 160일 이상 두어도 휴면타파가 되지 않았으므로 황산과 지베렐린과 같은 화학약품 처리에 의한 휴면타파 정도를 조사하였다(Table 2). 둥근잎유홍초 종자는 황산에 침지한 시간이 길어질수록 발아율은 증가하였고, 황산에 15, 20 및 30분 침지한 경우 63-75% 발아율을 보였다. 그러나 황산침지 20분과 30분 처리의 종자로부터 발아된 유식물체는 엽록소 형성 저해를 보였다(자료 미제시). 따라서 둥근잎유홍초 종자 휴면타파를 위한 황산침지 시간은 15분이 가장 적합한 것으로 판단되었다. 본 연구와 유사하게 미국실새삼(Buhler and Hoffman, 1999)과 헤어리베치(Kim et al., 2013) 종자의 경우 황산 침지시간이 길어질수록 이들 종자의 발아율도 증가하였다. 지베렐린 100 ppm에 24시간 침지한 경우 둥근잎유홍초 종자의 발아율은 단지 23%를 보여 무처리와 유의적인 차이가 없었다.
온도조건별 둥근잎유홍초의 발아율 및 생육
둥근잎유홍초의 적정 발아온도는 25°C에서 73%의 가장 높은 발아율을 보였고 지상부 및 지하부 생체중 가장 많았다(Table 3). 한편 15°C에는 25.6%의 낮은 발아율을 보임으로써 저온에서는 발아력이 감소하는 것을 확인 할 수 있었다. 그러나 35°C에서는 25°C와 유사한 발아율과 지하부 생체중을 보였으나 지상부 생체중은 유의적으로 감소하였다.
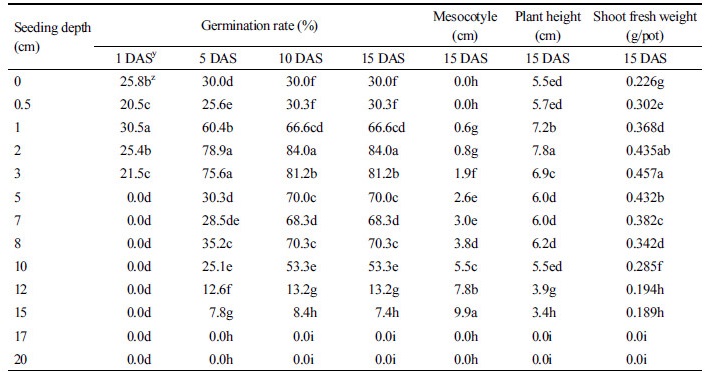
파종깊이별 둥근잎유홍초의 발아율 및 생육
둥근잎유홍초의 파종 심도별 발아율을 보면 2, 3, 5, 7, 8 cm 파종깊이에서는 발아율이 70-84%로 높았으나 12 cm와 15 cm 파종깊이에서는 각각 7%와 13% 발아율을 보였고, 17 cm 이상의 파종깊이에서는 발아하지 않았다(Table 4). 파종깊이 중 최고의 발아율은 1 cm에서 84%로 가장 높았다. 1년생 잡초는 보통 자연상태에서 토양 표층 1-2 cm 이내에서 발생하는데, 이것은 형성된 종자가 토양 표면에 낙하하였거나 또는 다른 지역으로부터 이동 ․ 전파되어 존재하게 되기 때문이다(Aldrich and Kremer, 1997). 중배축은 파종깊이 1 cm부터 발생하기 시작하여 15 cm까지 발생하였고, 파종깊이가 깊을수록 증가하였다(Fig. 1). 대부분의 잡초는 매우 얕은 토양층에서 발아가 잘되고 중배축이 발생하지 않으나, 파종심도가 깊을수록 중배축이 길어지고, 유묘가 토양표면까지 도달할 수 없으므로 사멸한다고 하였다(Raju and Steeves, 1998; Schafer and Chilcote, 1970; Wesson and Wareing, 1969). 그러나 본 연구에서 둥근잎유홍초 파종깊이 15 cm에서 중배축이 10 cm까지 발생하였고, 초장과 지상부 생체중도 대조구(파종깊이 0 cm)보다는 유의적으로 감소하였으나 건전한 생육을 보였다.

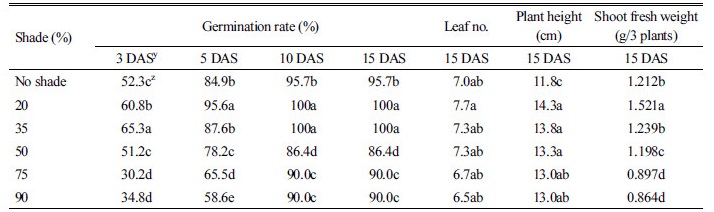
차광정도별 둥근잎유홍초의 발아율 및 생육
둥근잎유홍초는 차광 20%와 35%에서 100% 발아율을 보여 무차광(96%)에 비해 유의적으로 높았다. 또한 90% 차광에서도 90% 발아율을 보여 둥근잎유홍초 종자는 광조건과 상관없이 높은 발아율을 보였다(Table 5). 둥근잎유홍초의 엽수는 차광정도와 상관없이 무차광과 유의적인 차이가 없었다. 그러나 지상부생체중은 35% 미만의 차광 조건에서는 무차광 조건보다 높았지만, 그 이상으로 차광도가 높아지면 감소되는 경향이었다. 초장은 무차광에 비해 차광에서 유의적으로 증가하였으나 차광 정도간에는 유의적인 차이가 없었다. 일반적으로 차광은 식물체의 줄기신장에 영향을 준다고 알려져 있는데 잔대(Sicyos angulatus)에서는 차광율이 높을수록 초장 늘어난다고 보고 되었다(Moon and Pyo, 1981). 또한 가시박(Sicyos angulatus L.) 경우 60% 차광 조건에서는 무차광에 비해 초장이 3배 길게 나타났고, 지상부생체중도 2배 높았다(Oh et al., 2015). 그러나 Kim (2001)과 Hong et al. (2006)은 양지식물인 바위솔(Orostachys japonicas)의 경우 차광 정도가 높아질수록 지상부 및 지하부 생육이 낮아진다고 보고하였다.
토양수분별 둥근잎유홍초의 발아율 및 생육
발아 후 7일째 토양수분 10%와 100%(포화상태)에서는 발아율이 0%였고, 토양수분 30, 60, 80%에서는 60-83%의 발아율을 보였다(Table 6). 초장의 경우 토양수분 30, 60, 80%에서 각각 3.6, 4.5, 5.2 cm 였으며, 지상부 생체중도 토양수분이 30%에서 80%로 높아질수록 뚜렷하게 증가하였다. 따라서 둥근잎유홍초 종자는 토양수분이 아주 적거나 과습한 경우는 발아하지 않고 60-80% 수준에서 최적의 발아율과 생육을 나타내는 것으로 판단되었다.