서론
역사적으로 잔디는 자연선발, 비인위적 선발 또는 인위적 육종방법을 통한 선발 등 크게 세 가지 과정을 통해 현재의 자연 또는 인공적 환경에 적응하는 유전자원 풀(pool)이 유지되어 왔다고 볼 수 있다. 특히 잔디의 이용이 기존의 토양침식 방지 등 비교적 자연환경에 가까운 사방용에서 고도의 인공적 환경이 두드러진 스포츠용 등 다양한 환경 적응력을 보유한 잔디 유전자원이 필요해지면서 각국마다 자생 또는 도입 잔디 유전자원에 대한 중요성이 강조되고 있는 상황이다(Diesburg, 2000).
영명으로 한국잔디(Korean lawngrass)로 불려지는 zoysiagrass는 인도네시아를 비롯한 동남아시아의 열대 지역부터 한국, 일본, 중국 등 동북아시아의 온대기후 지역에 자생하는 것으로 알려져 있고, 분류학적으로 한국잔디 속의 식물은 현재 11개 종(species; Zoysia japonica, Z. macrantha, Z. macrostachya, Z. matrella, Z. minima, Z. pacifica, Z. pauciflora, Z. planifolia, Z. seslerioides, Z. sinica, Z. tenuifolia)이 적도를 기준으로 남북위 약 40도에 걸쳐 다양한 온도조건에 적응하면서 자연선발 과정을 거쳐 진화 및 적응해 온 것으로 알려졌다(Engelke and Anderson, 2003; Schwartz et al., 2010). 한국잔디는 지상포복경 및 지하경을 갖고 있어 낮게 자라며, 난지형 잔디 중에서 내건성, 내답압성, 낮은 비료요구성, 내한성 등 많은 장점을 가지고 있어 고품질 피복식물로 여겨져 왔다(Patton et al., 2017). 국내에서는 한국잔디의 이용범위가 더욱 다양해지고 사용면적도 확대되고 있기 때문에 다른 농작물과 달리 한국잔디의 재배농가 및 사용 면적은 과거 꾸준한 증가를 보이다가 현재는 감소하지 않고 유지되고 있는 것으로 보고되고 있다(Bae et al., 2013; Choi and Yang, 2006).
그러나 난지형 잔디 특성상 여름에 잘 자라지만 겨울에는 생육이 정지되며, 발아가 잘 되지 않아 주로 영양 번식에 의존하며 초기 피복율이 매우 느리고(Genovesi and Chandra, 2015; Patton et al., 2004), 겨울철 휴면으로 인한 탈색 및 갈색퍼짐병(large patch) 등의 병 발생 등 단점이 있다(Ma et al., 2013). 특히 품질이 양호하고 밀도가 높은 Z. matrella, Z. pacifica 등 일부 한국잔디는 내한성이 낮아 국내에는 제주 등 일부 지역에서만 생육이 가능하거나 특수 보온처리가 필요하여 이용 지역에 제한을 받기도 한다(Patton and Reicher, 2007; Patton et al., 2017). 도입육종을 통한 내한성이 증진된 Cavalier 품종(Engelke et al., 2002)의 육성, 종자발생이 우수한 우량 계통간 자연교잡을 통한 순환선발법(recurrent selection)을 기반으로 한 종자형 한국잔디 품종의 육성(Genovesi and Chandra, 2015), 금잔디(Z. matrella) 계통의 내음성이 우수하고 번식속도가 빠른 ‘Zeon’과 ’Zorro’(Patton, 2009), ‘Emerald’ 품종과 같이 들잔디와 비단잔디(Z. tenuifolia) 간 종간교잡을 통해 내한성과 번식속도를 개선한 품종 등 한국잔디류가 가지고 있는 단점을 극복한 성공적인 육종사례가 보고되고 있다(Volterrani et al., 2009).
하지만 보유하고 있는 유전자원이 빈약하거나 유전다양성이 확보되지 않으면 기존 사례와 같은 육종방법으로는 이러한 단점을 극복하는데 한계가 있다. 최근 형질전환 기술을 이용하여 기존 유전자원에서는 보이지 않던 새로운 형질의 품종개량 시도가 제초제 저항성, 내음성 및 내한성 증진, 칼라 잔디 육성 등에 시도되고 있다(Bae et al., 2008; Ganesan et al., 2012; Ge et al., 2006; Toyama et al., 2003). 본 연구의 목적은 돌연변이 육종의 하나로 방사선 에 의해 유도된 변이체의 형태적 및 생리적 변이 특성과 AFLP (amplified fragment length polymorphism) 마커를 활용하여 유전적 변이 확대를 정량적으로 평가함으로써 방사선과 같은 물리적 돌연변이원의 유전적 변이 창출과 형질개량 가능성을 알아보는 것이다. 향후에는돌연변이 육종기술을 이용하여 기후 변화를 대비하고 현재 국내외에서 활용할 수 있는 한국 잔디의 기후 한계선을 극복하고 변화하는 환경에 적응할 수 있는 한국잔디 계통 또는 품종육성을 위한 기초를 마련하는 것이다.
재료 및 방법
들잔디 종자의 감마선 및 양성자 처리
난지형 및 한지형 잔디 종류별 방사선 돌연변이 유도를 위한 선량 범위에 대한 연구는 본 연구팀이 관여하여 이미 보고한 바 있다(Lee et al., 2008). 한국 잔디의 경우 60Co에서 발생한 감마선을 건조 종자에 24시간 조사 처리할 경우 발아율 또는 실생묘 초기 생육에 차이가 있긴 하지만 50%의 발아 또는 생육 억제를 보이는 선량 수준이 약 250-300 Gy (1 Gy=100 rad) 정도로 알려졌다(Lee et al., 2008). 따라서 본 연구에서는 방사선 돌연변이를 유도하기 위하여 2010년 수입 들잔디 종자(덕창, 성남)에 한국 원자력 연구원 정읍 방사선 연구소에서 설치된 앞서 이용했던 동일한 60Co 선원을 이용하여 감마선량을 175, 200 그리고 250 Gy를 24시간 조사하여 실시하였다.
이온빔 조사는 2010년 한국원자력연구원에서 보유하고 있는 에너지 수준 45 Mev의 MC-50 사이클로트론의 양성자 발생장치를 이용하여 실시하였다. 양성자빔의 선량은 앞서 감마선 변이유도 선량을 목표로 175, 200, 그리고 250 Gy 발생에 맞게 빔전류와 조사시간을 조절하여 처리하였다. 감마선과 달리 양성자의 경우 높은 에너지 부여로 인한 생물변이 효과를 기대할 수 있으나 시료 투과율이 낮기때문에 직경 5.5 cm × 높이 1.0 cm 얇은 petri dish 에 종피처리가 완료된 들잔디 종자를 채운 후 밀봉하여 조사시료를 조제하였다.
들잔디 발아 개체의 온실 환경에서의 변이개체 및 형태적 특성 조사
감마선 및 양성자 처리 들잔디 종자는 원예용 상토로 채워진 사각형 트레이에서 발아시킨 후 발아된 개체로부터 형태적 변이 특성을 가진 개체를 선발하였고 특성을 조사하기 위해개체별로 직경 10 cm 크기의 플라스틱 화분에 이식한 후 비닐 온실에서 매일 오전 10시부터 시간별로 관수를 하며 관리하였다. 특별한 가온 장치 대신 태양광에 의존하여 관리하였고, 선발 기간동안(3-5월) 상토 제작시 포함된 초기 영양분이외에 추가 시비는 실시하지 않았다.

생육이 진행되는 동안 초기 실생묘 단계에서 양성자빔 및 감마선 처리구로부터무처리종자 개체와 비교하여 세엽형, 색소변이형, 다분지성, 왜성 및 저온 녹기유지 개체들을 선발하였다(Fig. 1). 평균 기온이 10℃ 이하로 떨어지는 시기에 감마선 처리구에서 선발됐던 녹기 지연, 색소 변이형, 왜성 변이체들은 기온이 올라가며 회복되는 동안 본래의 특성이 회복되어 선발대상에서 제외되었다. 반면에 선발 개체 중 변이특성을 보이는 개체들과 대조구 들잔디의 생육 특성을 분석하기 위해 20 cm×30 cm 화분에 이식한 후 잔디 생육 최적기인 6-9월 동안 중순에 월 1회씩 지상부 형태적 특성을 조사하였다.
생육 특성 항목으로 엽너비(leaf width; mm), 엽길이(leaf length; cm), 엽각(leaf angle; °), 초장(plant height; cm), 최하위엽 높이(height to 1st leaf from ground; mm), 포복경 굵기(stolon thickness; mm) 및 포복경 길이(stolon length; cm)를 측정하였다. 엽너비와 엽길이는 생육최성기 제1엽이 만개했을 때 그 중 제3엽의 크기를 조사하였고, 엽각은 수직분얼경의 중심축으로부터 제3엽이 벌어진 정도, 초장은 지표면으로부터 최상부까지의 길이, 최하위엽 높이는 지표면으로부터 첫 번째 잎까지의 높이, 포복경 굵기는 포복경 끝에서 2-3번째 마디 사이의 굵기, 그리고 포복경 길이는 포복경 끝에서 3-4번째 마디 사이의 길이를 각각 측정하였다. 모든 항목은 변이체 화분 당 5회 측정하여 평균값을 얻었고, 들잔디 변이체 간 평균값 비교는 Duncan’s multiple range test로 통계 비교하였다.
선발 개체의 포장 환경에서의 특성 조사
1차 온실 특성조사 후 다음 해에 외부 포장에서의 형태적 특성를 위해 충남대학교 농장 밭포장에서 완전임의배치법을 이용하여 대조품종으로 방사선 무처리 들잔디와 중지를 포함하여 선발 변이체 17 계통을 80 cm × 80 cm 크기를 기본 식재 면적으로 하여 3반복으로 실험포장을 설계하였고, 반복당 골프장 홀 커터를 이용하여 4개의 플러그씩 동일한 간격으로 식재하였다. 1차 온실 평가에서와 같이 2차 외부 포장에서의 생육 특성도 위에서 기술한 지상부 항목들과 조사 방법으로 측정하였다. 2차 포장 조사의 경우는 반복구 별로 임의로 5 개 식물체를 선정하여 측정값을 평균하여 반복을 대표하는 값을 구하였고, 3반복에서 얻어진 측정값은 SAS 프로그램을 이용하여 계통간 평균값을 비교하여 통계처리 하였다(SAS, 2001).
녹기 지연 들잔디 변이체의 세포막 안정 특성 검정
휴면기의 방사선 무처리 대조구와 녹기 지연 선발 변이체의 세포벽 안정 특성 차이를 알아 보기 위하여 휴면 도입 시기에 신선한 잎을 채취하여 전해질 유출 정도를 측정하였다(Lee et al., 2011). 비슷한 크기와 엽색을 나타내는 잎을 선발하여 길이 0.5 cm로 자른 후 증류수 10 mL을 넣은 50 mL 튜브에 0.1 g씩 침지시켜 3반복으로 준비하였다. 시료용액이 든 튜브를 32℃의 수조에 2시간 동안 방치 후 꺼내어 EC meter로 측정하였다(EC1). 측정한 샘플 튜브를 121℃에서 15분 동안 고압멸균 한 후 꺼내어 25℃에서 식힌 후 EC meter로 측정하였다(EC2). 전해질 유출의 정도는 EL=(EC1/EC2) × 100 공식에 따라 계산하였다.
세포막의 지질과산화 정도는 기존의 방법을 일부 응용하여 지질과산화를 통해 생성되는 malondialdehyde (MDA)를 측정하여 간접적으로 조사하였다(Reilly and Aust, 1999). 0.2g의 잎을 채취하여 마쇄한 뒤 10% (v/v)의 trichloacetic acid (TCA)와 thiovarvituric acid (TBA) 0.6% (v/v) 이 들어 있는 용액에 12,000×g 에서 15분간 원심 분리하여 상등액을 분리하고 95℃ 물이 들어있는 수조에서 15분간 중탕하고 빠르게 냉각시킨 후 다시 12,000×g에서 15분간 원심 분리한 뒤 최종 상등액만 취하여 532 nm에서 흡광도를 측정하여 600 nm의 비특정 흡광도를 제거한 뒤 계산하였으며 모든 측정은 3반복으로 수행하여 통계처리를 위하여 표준오차를 계산하였다.
잎의 엽록소 함량을 알아보기 위해 각 계통에서 유사한 발달 단계의 크기와 엽색이 비슷한 충분히 자란 잎을 선정하여 녹색을 띠는 잎의 중앙 부분을 SPAD-502 PLUS 엽록소 측정기(Spectrum Technologies, Aurora, IL, USA)를 이용하여 엽록소 함량을 나타내는 지표값(- 9.9-199.9)을 5반복씩 측정하여 평균값을 구한 뒤 통계처리를 위하여 표준오차를 계산하였다(Wang et al., 2012).
선발 변이계통간 유전다양성 평가를 위한 AFLP 분석
온실 및 포장 생육특성을 기초로 최종 선발된 세엽, 다분지 고밀도 및 녹기연장 변이 계통들의 유전적 다양성을 확인하기 위해 각 계통의 어린잎을 CTAB (Hexadecyltrimethylammonium Bromide) 방법(Doyle and Doyle, 1987)을 이용하여 genomic DNA를 추출하였다. AFLP 분석은 제한효소 EcoRI과 MseI으로 genomic DNA를 절단한 후 37℃에서 2시간 방치하여 특이적인 제한 절편의 말단에 EcoRI/MseI adapter를 연결하였다. 본 연구실의 기존 발표(Jeon et al., 2016)의 방법과 같이 절단된 DNA 절편은 1차 PCR 예비증폭(preamplication) 산물을 주형으로 하여 EcoRI에 형광표지한 총 11개 조합을 이용하여 2차 PCR 선택증폭(selective amplification)을 수행하였다(Fig. 3). 선택 프라이머를 이용한 2차 PCR은 touch-down PCR 방법(매회 0.7°C씩 온도 강하)으로 94°C에서 30초, 65°C에서 30초, 그리고 72°C에서 2분 동안 증폭 과정을 반복하여 총 12회 실시한 후, annealing 온도를 매회 0.7℃ 씩 낮추어 가며 추가로 30회 더 반복 진행하여 분석용 최종 산물을 확보하였다. 증폭된 최종 산물은 ABI 3130xl genetic analyzer (Applied Biosystems, CA, USA)를 이용하여 개체간 절편의 유무 및 크기 등 allele 정보를 확보하여 데이터 매트릭스를 작성하고, Molecular Evolutionary Genetics Analysis (MEGA; megasoftware.net) 프로그램의 neighbor-joining 방법을 이용하여 연관수형도(phylogenic tree)를 작성하였다. 마지막으로 Powermarker (v. 3.25; powermarker.software.inormer.com)프로그램을 이용하여 계통간 유전다양성 정도를 평가하기 위한 지표로 major allele frequency, Polymorphic information content (PIC) 그리고 genetic diversity 값을 결정하였다.
결과 및 고찰
선발 개체의 온실내 생육특성
초장, 엽장, 엽각, 포복경 길이는 변이체와 대조구 간에 유의한 차이가 없었으나, 질감의 기준이 되는 엽너비(mm)는 무처리 들잔디 야생형은 4 mm, 양성자빔 및 감마선을 조사한 잔디 변이체들은 약간 변화된 엽폭을 나타냈다(Table 1). Duncan의 다중검정(Duncan's multiple range test)으로 양성자빔 및 감마선을 조사한 잔디 변이체와 조사하지 않은 대조구의 유의성 검정을 한 결과 대조구(control)와 양성자빔 및 감마선으로 조사한 들잔디 선발 개체중에서 양성자빔을 250 Gy조사한 p250-6 계통만이 대조구보다 약 1 mm 정도 좁은 특성을 나타내 통계적으로 유의성있게 엽폭이 좁아진 것으로 나타났다 (Table 1).
또한 예지 저항성을 나타내는 지상부 첫번째 잎까지의 높이도 감마선과 양성자 처리를 받은 r200-14, r200-26, p200-14 등의 선발 계통에서 무처리 야생형에 비하여 지표면에 가까운 위치에서 발생하여 왜성형의 변이체 유도도 가능함을 보여주고 있다. 난지형 잔디의 경우 적절한 예초 높이는 뿌리의 발달을 촉진시키는 효과가 있지만(Wherley et al., 2011), 높은 관리수준으로 관리되는 스포츠용 잔디의 경우 고품질 잔디를 위해서는 낮은 예초 높이를 권장하므로 잎의 발생 위치는 들잔디 품종 육성에 중요한 요인이 될 수 있을 것이다(Frank et al., 2004).
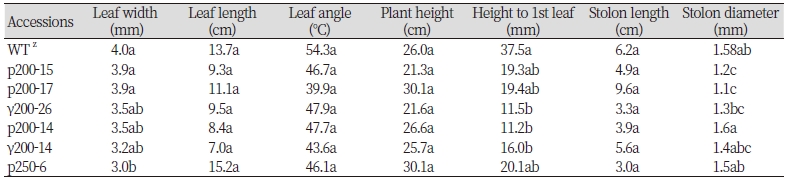
선발개체의 포장환경 생육특성
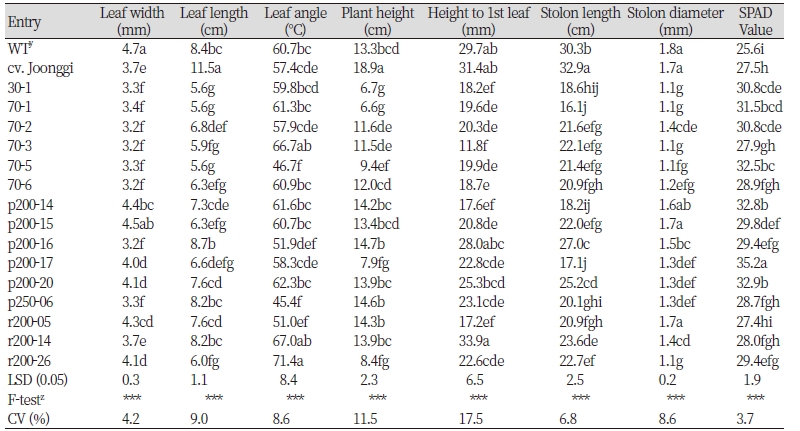
2차 외부 포장 환경에서의 선발 계통들의 생육 특성을 조사하기 위하여 1차 온실 평가에 활용된 6계통을 포함한 총 15 들잔디 변이계통을 무처리 들잔디 야생형과 중지 품종을 대조구로 하여 조사를 실시하였다. 세엽의 특징을 보이는 30-1, 70-1, 70-2, 70-3, 70-5, 70-6(3.2-3.4 mm)의 엽폭은 중지(3.7 mm)와 차이가 나지 않지만, 들잔디(4.7 mm)와 비교했을 때는 약 1 mm 이상 좁은 것으로 나왔고, 전체적으로 15 계통 모두 비교하여도 통계적으로 엽폭이 좁다는 것을 알 수 있었다(Table 2). 그리고 양성자빔을 처리한 p200-16(3.2 mm), p250-06(3.3 mm) 선발 개체의 엽폭도 들잔디보다 약 1 mm 이상 좁고, 중지와도 유사한 것으로 나와 세엽의 특징을 보였다. 1차 온실 평가에 포함된 6 계통의 엽폭은 포장 조사에서도 유사한 경향을 보이고 있었다. 특히, p250-6 선발 변이체는 온실 및 포장조사에서 무처리 야생형보다 약 1 mm 이상 좁은 특성을 나타내 질감이 섬세한 고품질 품종으로 상용화할 가능성이 높을 것으로 확인되었다.
한편 1차 온실평가때와는 달리 2차 외부 포장평가에서는 조사한 모든 항목에서 ANOVA 분석결과 계통간 유의적으로 차이가 발생함을 알 수 있었다. 1차 온실평가에서의 7계통보다 더 많은 선발 변이 계통이 포함되기도 하였고, 온실의 비교적 환경편차가 크지않은 상황과는 달리 외부에서의 온도 및 토양수분 등 생육에 영향을 미치는 요인들의 영향때문일 것으로 판단된다. 특히 계통간 유의적 차이를 보이지 않았던 초장, 엽장, 엽각, 포복경 길이 등에서도 들잔디 또는 중지 등 대조 자원들과 달리 왜성 특성을 보이는 계통들(30-1, 70-1, p200-17, r200-26), 엽각이 좁은 직립형 잎형태 계통들(70-5, p250-6)이 방사선 처리를 통해 유도되는 것을 알수 있었다(Table 2). 뗏장 생산에서 중요한 포복경의 길이는 대조 품종인 중지나 들잔디 만큼 빠른 생육을 보이는 변이 계통들이 선발되지는 않았다. 일반적으로 물리적 돌연변이원인 방사선의 경우 본 연구에서와 같이 고선량으로 처리되는 경우 생육 촉진보다는 억제되는 변이체의 발생이 많다는 기존의 밀, 보리, 양파 등 다른 작물과 유사한 결과를 확인할 수 있었다(Jan et al., 2011). 저선량과 달리 고선량에서의 포복경 생장속도 및 크기 등 생육억제의 원인으로 오옥신 합성, 핵산의 합성 감소와 함께 광합성 산물의 생성 및 타 조직으로의 분배가 감소되는 것과 관련이 있다고 알려지고 있다(Jan et al., 2012).
휴면기 녹기 지연 변이체의 세포막 안정특성 검정
휴면기와 같은 저온 조건은 광화학계를 손상시켜 엽록체에 과도한 양의 자유에너지가 발생되며, 자유도가 증가된 에너지는 엽록체 내에 풍부한 산소분자에 전달되어 초산소이온(O2-), 과산화수소(H2O2), 일중항산소(1O2)와 히드록시라디칼(.OH)과 같은 활성산소종(reactive oxygen species; ROS)을 생성한다. 활성산소는 원형질막과 엽록체를 비롯한 세포 내 여러 소기관들을 과산화 시키고 세포를 파괴하는 유해한 작용을 하며, 특히 생체막의 주요 성분인 불포화지질은 활성산소종의 공격에 취약하며, 과산화되면 MDA와 같은 과산화지질을 생성하고 전해질 누출이 증가되는 과산화적 증상이 나타나게 된다(Lee et al., 2011). 그러므로 녹기가 지연된 변이체와 대조구 간의 내한성 차이를 알아보고 저온에 대한 반응 차이를 밝히기 위하여 저온시기 휴면 상태에서 선발 변이개체들 중 녹기가 지연되는 특성을 보이는 계통들을 대상으로 지질과산화 정량분석, 전해질 유출을 통한 세포막의 물리적 안정성과 엽록소 함량을 조사하였다(Fig. 1).
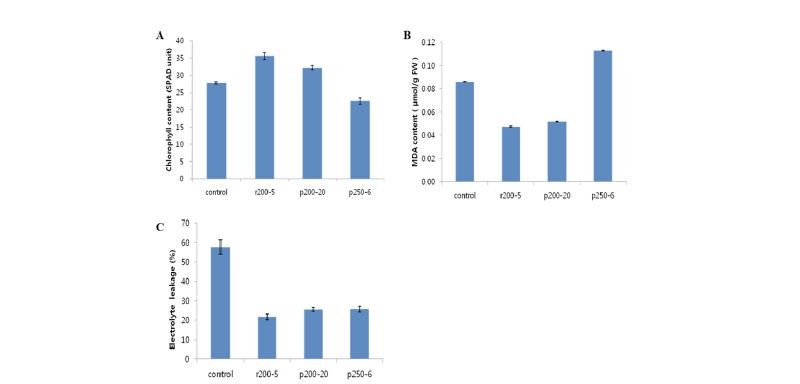
지상부 잎의 녹색이 유지되었던 2개의 변이 계통 r200-5와 p200-20에서 엽록소 함량이 야생형 들잔디 계통에 비하여 통계적으로 유의하게 높게 유지되고 있음을 알 수 있었다(Fig. 2). 지질의 분해 산물인 MDA 함량은 야생형 들잔디와 비교해서 상대적으로 잎의 녹기가 오래 유지되었던 r200-5와 p200-20 계통에서 50% 이상 낮게 나타났으나, 세엽 변이체 p250-6에서 가장 높게 나타나서 저온기 녹기 지연 특성은 없는 것으로 나타났다(Fig. 2). 전해질 유출(electrolyte leakage)은 예상대로 대조구에서 가장 높게 나타났고, 선발된 3개 변이체 모두에서 전해질 유출량이 대조구에 비해 감소하였으며 특히 r200-5에서 가장 낮게 나타났다. 엽록소의 함량을 나타내는 SPAD 지표 결과도 녹기 지연 2개 선발개체 r200-5와 p200-20 계통에서 통계적으로 유의하게 높게 나타났다. 세엽 변이 개체인 p250-6 계통은 안토시아닌 계통의 붉은 색이 많이 생성되었고, 지질산화 정도는 높지만 전해질 유출은 낮은 특성을 보여 저온 노출시 빠른 휴면 돌입으로 인한 높은 저온 적응성을 보인 반면, 녹기 지연과 내한성 기작은 상이하다는 추론을 해 볼 수 있다(Hara et al., 2003).
선발된 형태 돌연변이체의 AFLP 분석을 통한 유전적 다양성 확인
앞서 들잔디 선발 변이계통들의 지상부 형태 또는 저온 휴면기 산화스트레스에 대한 반응을 통하여 볼 때 방사선 처리에 의한 생리적 변화가 유전적 돌연변이와 관련 있음을 알 수 있었다. 이를 증명하기 위하여 제한효소 EcoRI 과 MseI을 조합을 통해 들잔디 유전체 상의 DNA의 특정 부위를 인식하여 절단함으로써 다양한 절편을 생성하게 하여 대립유전자의 다양성을 평가할 수 있는 AFLP 분석을 실시하였다(Fig. 3).
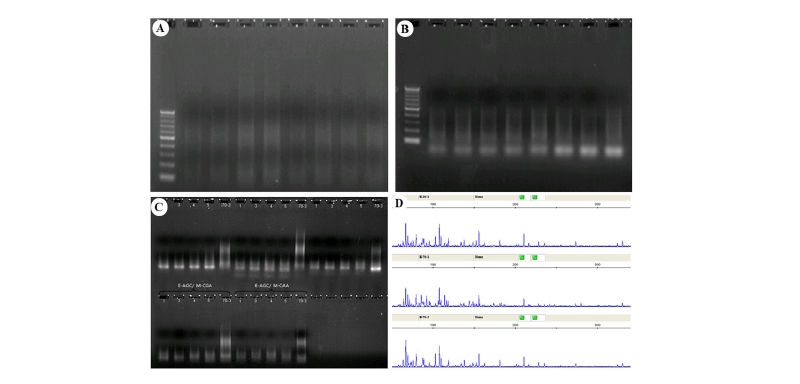
Fig. 3. Procedures to obtain different alleles at the amplified loci digested by the combinations of EcoRI and MseI restriction enzymes. (A) enzyme digestion, (B) preamplification, (C) selective amplification, and (D) chromatogram showing allele compositions of each sample from an ABI 3730x DNA sequencer.
제한효소는 총 11개 조합으로 구성하여 DNA 사슬에서 절편의 한 쪽 부분은 EcoRI에 의하여 5’-G/AATTC-3’ 염기배열을 인식하여 절단하고, 같은 절편의 반대쪽 부분은 MseI에 의하여 5’-T/TAA-3’ 염기배열을 인식하여 절단함으로써 다양한 길이의 동일한 절단 말단 특성을 보이는 DNA 절편만을 선별하여 allele 다양성을 분석하는 방법이다. Table 3에서 보여주는 것처럼 이 들 절편의 말단 부위에 3개의 염기로 구성된 어뎁터 종류를 달리 연결하게 되면 계통간의 특정 절편의 수, 즉 본 변이계통의 집단에서는 36-119개의 allele이 발생된다는 사실을 알 수 있었다. 이들 다양한 유전자 절편( fragment 또는 allele) 중 17개 들잔디 자원들간에는 평균 66개 중 55개의 유전자좌에서 차이를 나타내어 평균 78.5%의 다형성(polymorphism)을 보였다. 국내 자생 버뮤다그래스의 경우 동일한 제한효소 조합으로 절단했을 때 제한효소 조합당 평균 78개의 절편이 얻어진 것과 비교해 볼 때 유전 다양성 평가를 위해 충분한 비율이라고 할 수 있다(Kang et al., 2008). 본 연구에서 얻어진 78.5%의 다형성 유전자의 비율은 화본과의 다른 식물인 숲개밀(Brachypodium spp.)의 자연 및 인공 자원으로 구성된 2개의 집단을 비교한 결과에서 방사선 처리에 의하여 59%에서 79%로 다형성 유전자의 비율이 높아진 것과 같이 방사선 처리가 유전다양성 확대에 기여하고 있음이 AFLP 분석을 통하여 재증명되게 되었다(Zhang et al., 2012).
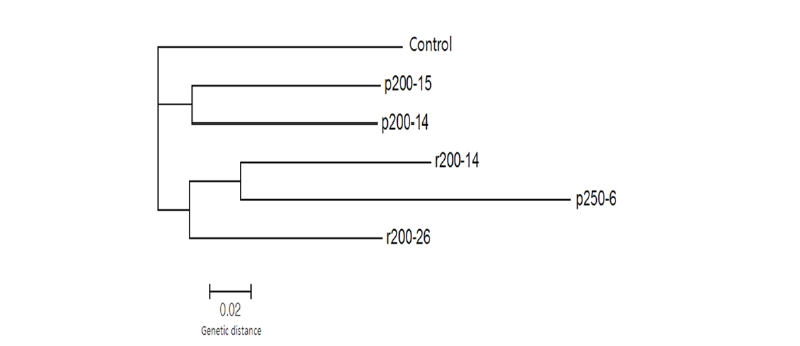
또한 17개 계통간에 최다 빈도를 나타내는 major allele 비율은 0.85였고, PIC는 0.16으로 나타나서 전체 유전자 중 약 16%가 감마선 또는 양성자에 의하여 유전적인 변화가 발생되었다는 것을 간접적으로 판단해 볼 수 있었다. AFLP 분석을 통해 생성된 다형성 유전자들을 기본으로 유전적 유사 또는 차이를 표시하는 계통도를 작성하여 유연관계를 비교한 결과, 대조구 야생형 들잔디는 다른 방사선 처리에 의해 유도된 변이 계통들과 유전적으로 제일 거리가 멀다는 것을 알수 있었다(Fig. 4). 즉 선발된 변이개체들은 들잔디 야생형과 유전적으로 다른 group에 속하고 있었고, 왜성 특성을 보이며 잎의 각도가 수직에 가까운 r200-26 변이체가 독립된 한 그룹을 만들고, 양성자빔을 이용한 p200-14와 p200-15가 유사하게 동일한 그룹을 형성하고 있고, 엽폭이 세엽인 특성을 보이는 r200-14와 p250-6이 같은 유전 그룹에 형성되었다. 그 중 양성자 처리에 의하여 유도된 p250-6 선발 계통이 다른 비교 계통들에 비하여 유전적 유사도가 가장 낮게 나타나 유전자의 변이가 제일 많이 발생하였다는 것을 추론할 수 있었다(Fig. 4).
|
Table 3. Morphological diversity of radiation-irradiated zoysiagrasses grown under greenhouse conditions. 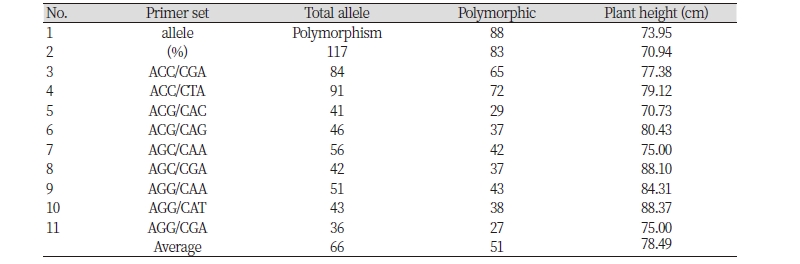
|
본 연구를 통하여 방사선 처리 시 들잔디의 경우 왜성, 분지성 또는 세엽형 변이체 등 다양한 형태적 차이를 유발하고 있음을 확인하였다. 하지만 유전적 변이가 제일 많이 발생한 p250-6 변이 계통의 경우 세엽 표현형이 많은 유전자에 의하여 조절되는지 또는 소수의 방사선 민감 유전자에 높은 빈도의 변화가 발생한 것인지 등 각각의 형질과 연관된 유전자의 변이 특성과 함께 이들 유전자들을 조절하는 유전 인자들(예, miRNA)의 작용 특성도 연구되어야 돌연변이 개체들의 표현형과 발생 기작을 더 구체적으로 이해할 수 있을 것으로 판단된다 (Subburaj et al., 2017).

