서론
인류 삶의 질 향상, 기후변화, 자연 보전 및 생태환경보호, 생물 자원화 등에 대한 관심이 증가됨에 따라 전 세계적으로 생물다양성 보전과 유지를 위한 노력이 매우 활발히 진행되고 있는 상황이다. 우리나라에서도 이를 위한 실천방안중의 하나로서 생태계 위해우려종및 생태계교란 생물종을 확대 지정함과 동시에 이들의 모니터링, 제거활동, 생물다양성 위협 외래생물 관리 R&D 사업 추진 등 보다 체계적인 관리를 시행하고 있다(Kim, 2016). 2016년 국내에서 추가된 두 가지의 생태계교란 식물종은 Spartina속의 외래식물인 갯줄풀(Spartinaalterniflora, Smooth cordgrass)과 영국갯끈풀(Spartinaanglica, Common cordgrass)이다. 이들의 국내 서식은 2008년경부터 시작된 것으로 추정되지만 2015년도에 처음으로 공식 보고되었으며(Kim et al., 2015), 현재 초기 발생지인 강화도와 전남 진도에서 가장 많은 분포로 서식되고 있고, 영종도, 웅진군 신도, 안산시 대부도, 충남 서천군 등에서도 발생이 확인되어 지역적으로 점차 확산되고 있는 추세이다.
Spartina속 식물은 전세계적으로 약 17종이 알려져 있으며(Hedge et al., 2003; Peterson et al., 2007), 이들 중 S. alterniflora, S. anglica, S. densiflora 등과 같은 몇가지 종은 빠르게 생장 번식하면서 주변의 식생을 파괴하고 생물다양성을 감소시키며 갯벌 및 염습지의 기능을 상실시키기 때문에 전세계적으로 생태환경 교란 침입성 식물로 지정하여 여러가지 방법으로(물리적, 화학적, 생태학적, 종합적 방제방법) 이들의 관리에 많은 노력을 기울이고 있고, 특히 미국, 영국, 호주, 중국 등에서의 연구사례가 많이 보고되고 있다 (Kim, 2016).
침입성이 강한 외래식물은 적응, 생장, 번식력이 상대적으로 우월한 특성을 지니고 있다. 따라서 새로운 지역에 유입되면 빠른시간내에 우점화되면서 기존의 식생을 파괴하면서 생물다양성을 감소시키고 아울러 부차적인 여러 가지 환경문제를 일으키기 때문에 관리의 적정시기를 놓치면 이를 제어하기 위한 사회적 비용이 기하급수적으로 증가하게 된다(예, 국내에서의 가시박 발생 및 확산문제). 무엇보다 이러한 현상은 자연계에서 일어나는 일이기 때문에 발생초기에 신속히 대처하는 것이 가장 바람직 할 것이다. 해양수산부 보도자료에 의하면 우리나라에 출현한 S. anglica와 S. alterniflora의 2018년도 현재 발생면적은 31,333 m2 정도라고 한다. 이는 그다지 높은 수준이 아니기 때문에 경제적 방제의 골든타임을 놓친 것 같지는 않지만 Spartina속 식물은 왕성한 번식특성을 가지며 불량환경에 대한 적응력이 뛰어나고, 유묘상태에서 갈대를 비롯한 여러 화본과 초종과의 식별이 쉽지 않은 점 등을 고려해 볼 때 잠시 방치하면 합리적 관리 한계를 손쉽게 벗어날 우려가 높다(Kim, 2016). 그리고 국내에 유입, 적응하는 과정에서 생리생태적으로 어떠한 특성변화가 유도되었는지도 모른다. 따라서 본 연구는 국내 Spartina속 식물의 기본 생태특성을 조사하여 이들의 방제 및 확산방지를 위한 기초자료를 얻기 위하여 제반 실험을 수행하였다.
재료 및 방법
종자 및 근경 수집
갯줄풀(Spartinaalterniflora, SPAAL) 종자(영과, caryopsis)의 경우, 10월 중순에 전남 진도에서 수집한 이삭(spike)을 약 1달간 온실 건조시킨 후 가볍게 털어 얻어진 비교적 충실한 종자를 선별하여 사용하였다. 영국갯끈풀 (Spartinaanglica, SPAAN) 종자(영과, caryopsis)는 11월 중순에 강화도에서 수집한 이삭을 약 10일간 온실 건조시킨 후 가볍게 털어 얻어진 비교적 충실한 종자를 선별하여 사용하였다. 이들의 수집 종자는 실험전에 실온조건에서 스포탁 유제(한국삼공주식회사, 한국) 2000배 희석액에 하룻동안 침지하여 소독처리하였다. 영국갯끈풀 근경(rhizome)은 11월 중순에 강화도에서 수집하여 정선한 다음, 습기있는 비닐봉지에 넣고 4℃ 냉장상태에 보관하였다.
저장조건별 종자(영과)발아
종자의 휴면타파 및 발아조건을 알아보기 위하여 4가지 저장조건, 즉 건조한 상태(I), 수돗물 침지(II), 1% KNO3 용액 침지(III), 인공해수(염도 0.79%) 침지(IV) 상태로 종자를 저온(4℃) 냉장고에 보관하면서 경시적으로 발아실험을 수행하였다. 인공해수는 수돗물에 해수염(Instant Ocean Sea Salt, United Pet Group, Inc. Cincinnati, OH 45255)이 1% 되도록 조제하여 사용하였고, 인공해수의 염도를 염도계 (PAL-SALT, ATAGO, Japan)로 조사하였다.
발아실험을 위해서 직경 9.5 cm petri-dish에 여과지 1매를 깔로 수돗물 15 mL을 분주한 다음, 쭉정이를 최대한 배제시킨 정선된 종자를 50립씩 3반복 치상하였다. 이를 낮 25℃/밤 19℃ (12 h 광주기)의 생육실에 30일까지 보관, 증발된 수분량은 수돗물로 수시로 보충하면서 발아율을 조사하였고, 종자로부터 뿌리 또는 신초가 0.1 mm 이상 출현된 것을 발아된 것으로 간주하였다.
휴면타파된 종자의 재건조에 따른 2차휴면 유도여부 조사
수돗물에 침지된 상태로 저온냉장고(4℃)에 4개월 두어 휴면이 거의 타파된 갯줄풀 종자를 사용하였다. 이들을 3가지 온도조건에서 건조처리 하였는 바, 4℃ 저온건조는 냉장고에, 25℃ 저온건조는 생육실에, 40℃ 고온건조는 열풍건조기에 각각 4일, 9일동안 두고 건조처리로 인한 종자 생체중 감소율을 측정하였다. 건조처리된 종자의 발아실험을 위해서 직경 9.5 cm petri-dish에 여과지 1매를 깔로 수돗물 15 mL을 분주한 다음, 종자를 50립씩 3반복 치상하고, 이를 낮 25℃/ 밤 19℃ (12 h 광주기)의 생육실에 13일까지 보관하면서 발아율을 조사하였다. 그 후 발아되지 않은 것들을 모아 수돗물에 침지하여 다시 저온조건에 110일동안 보관한 다음, 앞서 수행했던 발아실험과 동일조건으로 재발아 여부를 조사하였다.
암(dark), 온도 및 염도가 종자발아에 미치는 영향
명암조건이 갯줄풀 발아에 미치는 영향을 조사하기 위하여, 1% KNO3 용액에 4개월 이상 저온침지하여 휴면이 타파된 갯줄풀 종자를 대상으로 10일간 명조건(light), 5일 암/5일 명조건, 10일 암조건(dark)에 각각 치상하여 발아정도를 조사하였다. 기타 발아실험 조건은 전과 동일하였다.
온도조건이 갯줄풀 발아에 어느정도 영향을 미치는지 알아보기 위하여, 1% KNO3 용액에 4개월 이상 저온침지하여 휴면이 타파된 갯줄풀 종자를 대상으로 낮 25℃/밤 19℃ (12 h 광주기)과 낮 35℃/밤 25℃ (12 h 광주기)의 두 조건에서 10일간의 발아율 시험을 실시하였고 이후 발아된 개체들의 유묘 생장상태도 조사하였다.
염도가 갯줄풀 발아에 미치는 영향을 조사하였다. 시중에서 구입한 천일염을수돗물에 녹여 0-6%의 염도가 되도록 한 다음 이를 시험용액으로 하였다. 휴면이 완전히 타파된 갯줄풀 종자를 여러 염도의 용액에 50립씩 3반복 치상한 다음, 낮 25℃/밤 19℃ (12 h 광주기) 조건에서 15일간 두어 10일째에 초기 유묘 생장상태를, 15일 째에는 최종 누적 발아율을 조사하였다. 이후 발아되지 않은 종자들에 대해서는 수돗물에 재치상하여 10일 동안 두면서 발아력 회복 여부를 추가로 조사하였다. 기타 발아실험 조건은 전과 동일하였다.
영국갯끈풀 근경생장에 미치는 염도의 영향
수돗물에 천일염을 녹여서 0-10% 범위내에서 여러 염도의 인공해수를 준비하였다. 사각폿트에(33 cm×20 cm×8.5cm) 건조 공시토양 (논흙+부농 수도용 상토, 3:2, v/v) 4 L를 담고, 신초가 2-4개 붙어있는 영국갯끈풀 근경(rhizome) 절편을 폿트당 3개씩 심었다. 재식한 폿트를 여러 염도의 물 6.5 L가 담긴 사각수조에 담궈 충분히 흡수시켜 표토까지 물에 잠기도록 하고 이를 온실 무가온상태의 베드에 옮겨 생육시켰으며, 수분 증발양 만큼 수돗물을 매일 공급하여 초기염도가 꾸준히 유지되도록 관리하였다. 한편 대조구로서 일반 밭조건과 같은 상태로 수돗물만 공급하는 처리구도 설정하였다. 재식후 53일째에 생육조사를 하였는 바, 지표로부터 5 cm 부위를 잘라 지상부 신초 길이와 생체중을 조사한 후, 이를 60℃에 3일동안 열풍 건조시킨후 건물중을 측정하였다. 지하부가 남아있는 폿트는 수돗물로 5회 이상 세척한 후, 초기 2-3일 동안 1x mDM 영양배지를 지표면으로 공급한 후 온실에 그대로 6주 재배하여 성장된 신초길이를 측정함으로써 재생여부를 평가하였다.
결과
갯줄풀 및 영국갯끈풀의 종자형태와 임실율
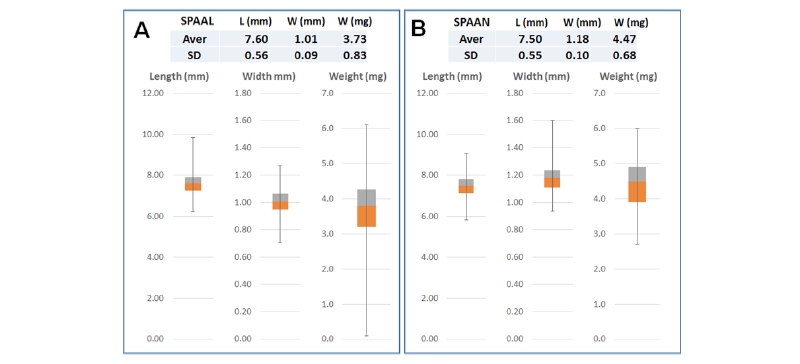
국내의 진도에 서식하고 있는 갯줄풀과 강화도 등에 발생하고 있는 영국갯끈풀의 실제 임실정도(영과중에서 성숙된 종실의 비율)는 다년생 식물임에도 불구하고 두 종 공히 80-85% 정도로 높은 편이었다. 종실(grain) 평균길이의 경우 갯줄풀이 7.6 mm로서 영국갯끈풀보다 약간 높았고, 종실 폭의 경우는 이와 반대로 갯줄풀이 1.01 mm로서 상대적으로 낮은 경향을 보였다. 종실의 개당 평균 건물중에 있어서는 갯줄풀(3.73 mg)보다 영국갯끈풀(4.73 mg)이 높았다(Fig. 1).
갯줄풀 및 영국갯끈풀 종자의 휴면타파와 발아특성
Cordgrass 종자(영과)는 종(species) 및 생장조건, 저장상태에 따라 휴면을 달리 가지는 것으로 알려져 있다(Biber and Caldwell, 2008; Marks and Truscott, 1985). 본 실험에서는 국내에 서식중인 cordgrass 종자의 4가지 저장조건별 휴면타파 여부와 발아력을 알아본 결과, 갯줄풀(SPAAL)의 저온건조(I)는 전혀 발아되지 않았고 나머지 3가지 침지 저장조건에서는 40일 저장 이후부터 발아율이 증가하기 시작하여 100일 이상 저장시 발아율이 80% 내외로서 휴면타파가 완성되는 경향을 보였다. 휴면타파가 잘 이루어졌던 이들 3가지 저장조건 중에서도 1% KNO3 (III) < 담수(II) < 인공해수 용액 침지(IV) 순으로 양호한 발아패턴을 보였다(Fig. 2A). 한편 영국갯끈풀의 경우는 약간의 다른 반응을 보였다. 즉 수확후 건조된 상태의 것은 13% 정도의 발아율을 보이다가 저온건조 저장기간이 진행될수록 발아율이 낮아지는 경향을 보였다(Fig. 2B). 한편 침지 저장의 경우는 저온저장 20일째에 40%정도 발아율을 나타내다가 저장 100일 이상에서 52-65%의 발아율이 보여 갯줄풀 종자보다는 낮은 발아력을 보였다. 침지저장 3가지 조건 중에서도 갯줄풀 종자에서와는 달리 수돗물 담수저장(II)에서 상대적으로 낮은 발아율을 보이고 1% KNO3 용액(III)과 인공해수 용액 침지(IV)는 비슷하게 상대적으로 보다 양호한 발아패턴을 보였다(Fig. 2B). 전반적으로 두종 공히 해수상태의 저온 침지가 가장 우수한 휴면타파 효과를 보였으며, 완전한 휴면타파에 요구되는 침지기간은 100일 이상이었다.
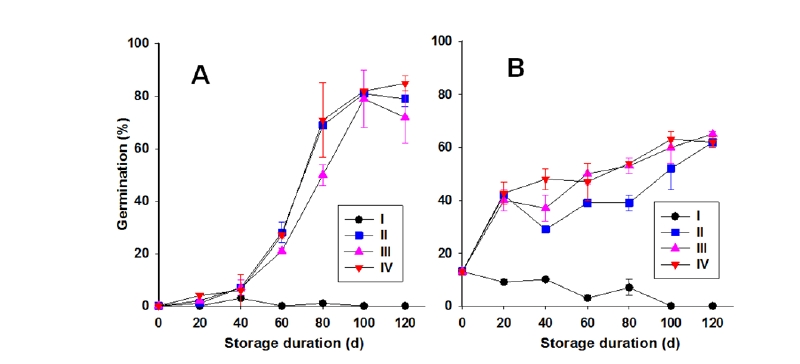
Fig. 2.Dormancy breaking of S. alterniflora (A) and S. angulata (B) seeds according to different storage conditions. I: Drying at 4℃, II: Soaking in tap-water at 4℃, III: Soaking in 1% KNO3 solution at 4℃, IV: Soaking in artificial seawater of 0.8% salinity at 4℃. Vertical bars indicate standard deviation of mean value of three replicates.
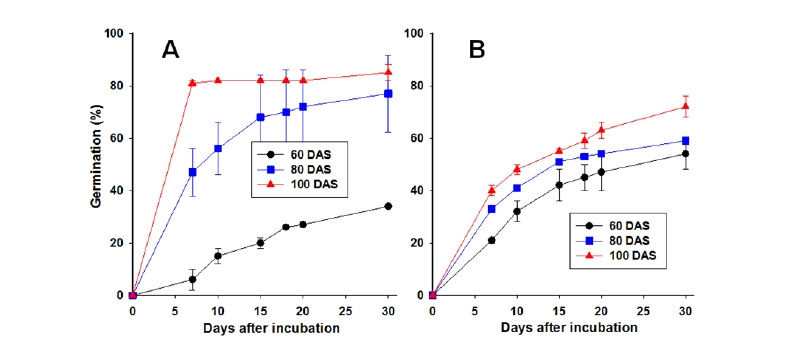
한편, 두 종의 발아속도 차이를 알아보고자 인공해수에 60, 80, 100일 저온침지된 종자의 배양기간별 누적발아율을 조사하여 발아속도를 비교해 본 결과는 Fig. 3에서와 같다. 갯줄풀의 경우는 침지저장 기간이 길어질수록 발아율이 높아짐과 동시에 발아속도가 빨라져 100일 저장된 것은 치상후 7일만에 80% 발아가 완성되었다(Fig. 3A). 그러나 영국갯끈풀의 경우는 침지저장 기간이 길어질수록 발아율은 약간씩 높아지지만 발아속도는 큰 변화가 없어 100일 저장시에도 치상후 7일째 발아율이 40%, 30일째 발아율이 70% 내외 정도를 나타내었다(Fig. 3A). 이는 영국갯끈풀이 갯줄풀보다 종자발아력이 상대적으로 낮음과 동시에 매우 완만한 속도로 발아하는 특성을 보유함을 보여준다.
휴면타파된 종자의 재건조에 따른 2차휴면 유도여부
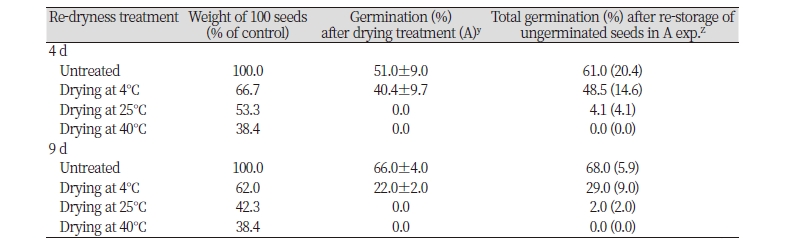
일부 식물종자는 휴면타파후 발아력을 보유하고 있지만 건조와 같은 불량환경에 접하면 2차휴면에 돌입하여 생존을 유지하는 특성이 있다(Née et al., 2017). 휴면타파된 갯줄풀 종자가 이처럼 다시 건조 되었을 때 2차 휴면에 들어가는지 아니면 바로 고사되는지를 알아보고자 실험하였다. 그 결과, 4일동안 4, 25, 40℃에 건조시켜 초기 생체중의 33.3, 46.7, 61.6%가 감소된 경우, 치상 10일째의 발아율이 무처리구의 51.0%보다 낮은 40.4, 0, 0%의 발아율을 각각 나타내었다. 여기에서 미발아된 종자를 모아서 100일 이상동안 저온침지시킨 다음 재발아되는지를 확인한 결과, 무처리구의 발아율이 20.4%인 반면 4, 25, 40℃ 건조처리구의 경우엔 14.6, 4.1, 0.0%를 보여 매우 낮은 수준을 나타내었다(Table 1). 이를 재확인하기 위하여 다른 실험을 통해 9일동안 4, 25, 40℃에 건조시켜 초기 생체중의 38.0, 57.6, 61.6%가 감소된 경우 치상 10일째의 발아율이 무처리구의 66.0%보다 낮은 22.0, 0, 0%의 발아율을 나타내었다. 여기에서 미발아된 종자를 모아서 100일 이상동안 저온침지시킨 다음 재발아되는지를 확인한 결과, 무처리구의 발아율이 5.9%인 반면 4, 25, 40℃ 건조처리구의 경우엔 9.0, 2.0, 0.0%를 보여 매우 낮은 수준의 발아회복을 나타내었다(Table 1). 만일 건조처리에 의해 2차휴면에 돌입되었다면 휴면타파처리로 인해 적어도 4, 25℃ 건조처리구에서는 무처리구에 근접한 발아율을 보였어야 할 것이다. 45℃ 처리구에서 발아가 전혀 이루어지지 않았던 것은 건조와 함께 고온으로 인해 종자가 사멸되었기 때문인 것 같다. 이상의 결과를 종합적으로 검토해 볼 때, 갯줄풀의 경우 휴면타파된 종자가 건조처리로 인해 재휴면에 돌입되지는 않는 것 같다고 여겨진다.
암(dark), 온도 및 염도가 종자발아에 미치는 영향
명암조건이 갯줄풀 종자발아에 미치는 영향을 조사하기 위하여, 휴면이 타파된 종자를 대상으로 10일간 명조건(light), 5일 암/5일 명조건, 10일 암조건(dark)에 각각 치상하여 발아정도를 조사한 결과(Fig. 4), 암조건에서도 발아력의 큰 감퇴없이 정상적으로 발아되는 특성을 보였다(Yuan and Shi, 2009).
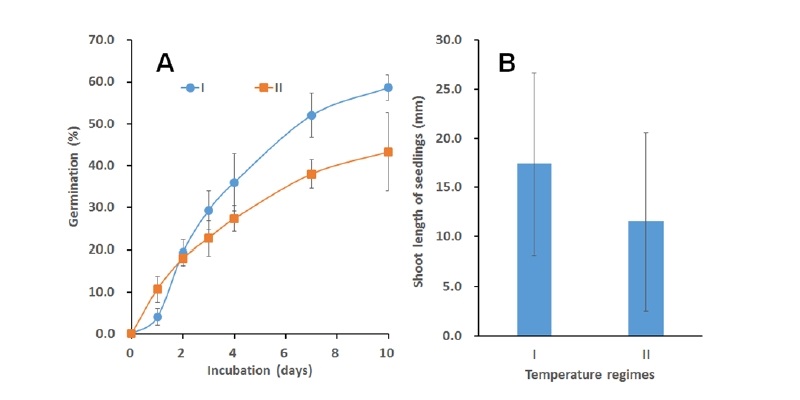
갯줄풀의 발아는 25℃ 및 낮 26℃/밤 16℃(14 h 광주기) 에서 양호한 것으로 보고되었으나(Yuan and Shi, 2009) 우리나라의 여름기온조건에서도 발아가 가능할지를 알아보기 위하여 비교적 고온조건(낮 35℃/밤 25℃, 12 h 광주기)에서의 발아력과 이후 발아된 개체들의 유묘 생장상태를 조사하였다. 그 결과, Fig. 5에서 보는 바와 같이 보통의 발아조건(낮 25℃/밤 19℃, 12 h 광주기) (I)에서는 60% 정도의 발아율을 보인 종자들이 고온조건(낮 35℃/밤 25℃, 12 h 광주기) (II)에서는 42% 정도의 발아율로 낮아졌고 이들의 초기생장도 보통의 발아온도 조건보다 33.3%가 감소된 생육상태를 보였다. 이는 여름의 고온 조건에서도 상대적으로 낮을지라도 갯줄풀 종자가 발아되어 생육될 수 있음을 시사해준다.
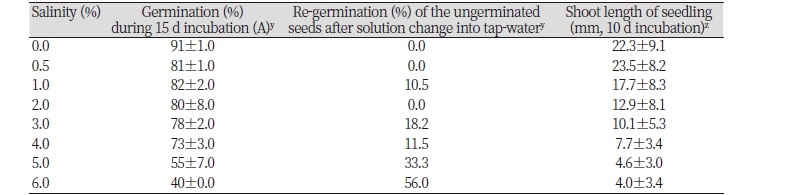
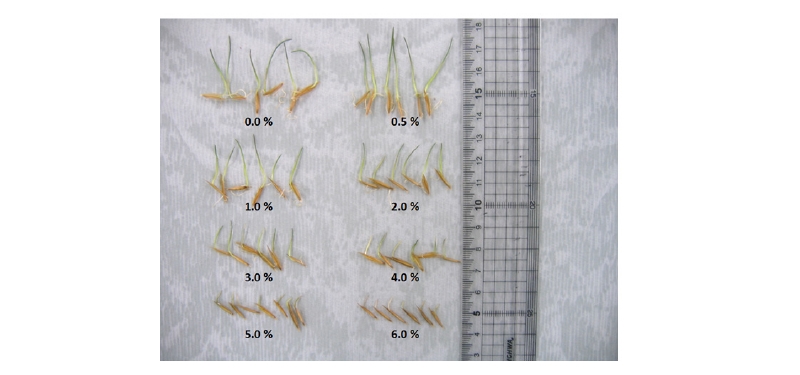
한편 갯줄풀 종자의 발아 및 초기생육에 염도가 어느 정도 영향을 미치는지 알아보고자 수행한 실험의 경우, 담수조건에의 발아율이 91%로서 염도가 0.5% 이상인 인공해수조건보다 발아율이 높은 경향이었다. 그러나 0.5-6% 범위의 염도에서는 염도가 높을수록 발아율이 억제되는 경향으로서 6% 염도에서는 40%의 발아율을 나타내었다(Table 2). 그런데 본 염도조건에서 발아되지 않았던 종자를 회수하여 세척한 다음 수돗물에 재치상하여 발아가 회복되는지를 조사한 결과, 3-6% 염도처리구에서 발아가 정상적으로 회복되는 개체들이 많았다. 즉, 6%의 고염도 처리를 예로 들면, 본 조건에서 15일간 침지되어 있을지라도 대부분의 종자는 죽지 않은 상태에 있다가 염도가 낮은 정상 환경에 옮기면 발아가 회복되며 초기 치상한 종자들의 약 20% 정도만 6% 고염도 처리에 의해서 죽는 것으로 여겨졌다. 한편 발아후 생장한 개체들의 생육상태를 조사해 보았을 때, 무염조건에서도 매우 양호한 생장을 보여 절대적 염생식물(obligate halophytes)은 아니었다. 그리고 0.5% 저염도에서는 유묘생장이 약간 높았으나(통계적 유의차는 없음) 그 이상의 염도에서는 염도가 높을수록 초기생장이 낮았고, 특히 4% 이상의 고염도에서는 생장억제가 심한 편이었다(Table 2, Fig. 6). 이들의 결과로 보아 갯줄풀 종자는 담수 뿐만 아니라 기수(brackish water) 및 해수에서도 발아하여 생존할 정도의 내염성이 있는 것으로 판단되었다.
타 연구자들의 결과에서도 이와 유사하게 갯줄풀 종자의 염도(NaCl)에 대한 발아한계 최고농도는 6-8%이었으며 생장반응은 0.5% NaCl용액에서 가장 좋았다 (Mooring et al., 1971). 또한 갯줄풀 종자는 40 g NaCl L-1의 고염도에서 (Wijte and Gallagher, 1996) 또는 200 mM 이상의 염도에서 (Li et al., 2010; Marks and Truscott, 1985) 발아율이 감소되었다는 보고가 있다.
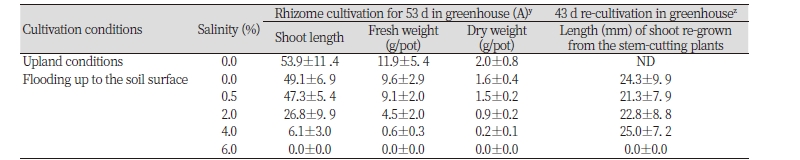
영국갯끈풀 근경생장에 미치는 염도의 영향
Cordgrass의 근경(rhizome)이 염도에 대해 어떠한 생장반응을 나타내는지 알아보기 위하여 영국갯끈풀을 대상으로 조사해 보았다. 그 결과, 근경 재식후 지하부가 물에 담겨진 상태로 자랄 때보다 밭조건 상태에서 자랄 때 더 양호한 생육을 보여 건물중 기준으로 약 25%의 생장 증가가 있었다 (Table 3). 지표면까지 물을 채워 담수상태에서 재배할 경우, 염도가 높을수록 생육이 억제되며 4% 염도 이상에서는 거의 자라지 못하고, 6% 이상에서는 모두 고사되는 경향이었다(Table 3, Fig. 7). 한편 약 50일동안 여러 농도의 염도에 생육하다가 염도가 없는 조건으로 전환하였을 때 4% 이하의 염도에서는 근경 모두가 정상 회복되어 생장을 했지만 6% 이상에서는 회복되지 못했다(Table 3). 이들의 결과로 보아 갯줄풀 종자에서처럼 영국갯끈풀 근경도 담수 뿐만 아니라 기수(brackish water) 및 일반 해수에서도 출아(shooting)하여 생존할 정도의 내염성이 있는 것으로 판단되었고, 얕은 담수(침지) 조건보다 밭조건에서 더 잘 자라는 특성을 보였다. 한편 고농도의 천일염을 사용할 경우 cordgrass의 방제가 가능함을 시사해 주지만 어느정도의 처리시간을 두어야 할 지는 추후 연구가 필요하다. Fuchen and Fang (2007) 연구에서도 100 mmol L-1 미만의 낮은 염도에서는 갯줄풀 생장이 촉진되었지만 100 mmol L-1 이상의 고염도에서는 생장이 억제된다고 보고하였다.
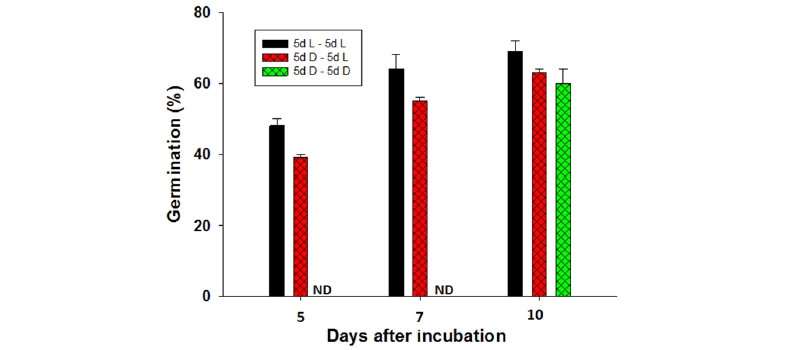
Fig. 4.Germination of S. alterniflora seeds in the different light-dark regimes. L and D represents light and dark, respectively. ND: In the case of 5d D - 5d D, germination was not determined at 5 d and 7 d after incubation. Vertical bars indicate standard deviation of mean value of three replicates.

고찰
어떤 지역에서의 침입식물 확산에 영향을 미치는 가장 핵심적인 열쇠는 종자의 착립율(seed set) 또는 종자번식(seed propagation) 능력이다(Daehler, 1998; Xiao et al., 2009). 갯줄풀(S. alterniflora)과 영국갯끈풀(S. anglica)은 다년생이라서 근경으로도 번식하지만 종자로부터 발아되어 유묘(seedling)가 정착한 이후 당해년도 또는 이듬해가 되면 새로운 이삭이 생긴다. 따라서 종자 임실율이 높고 종자 발아력이 우수하면 보다 빠른 속도로 확산될 것이다. 보고된 자료의 조사에 의하면 cordgrass 식물의 종자 임실율이 종(species)과 서식지에 따라 5%에서 85%에 이르기까지 다르다(Kim, 2016). 본 연구에 의하면 국내에 서식중인 갯줄풀과 영국갯끈풀은 종자의 임실율이 공히 80-85%로서 꽤 높은 편이었으며 탈립도 잘 되는 편이었다. 그러나 발아특성면에서는 두 종간에 약간의 다른 특성을 나타냈다. 즉, 휴면타파된 성숙한 영과(caryopsis)의 경우 갯줄풀은 70-90%, 영국갯끈풀은 60% 내외의 발아율을 나타내어 상대적으로 갯줄풀이 높았다. 그리고 이삭으로부터 수집한 성숙한 갯줄풀 영과는 발아력을 갖고있지 않아 성숙중에 보다 빠르게 휴면상태에 접어드는 것으로 여겨졌지만(데이터 제시 생략), 영국갯끈풀은 휴면으로의 점입속도가 완만하여 수확당시의 종자발아력이 10-15% 유지되다가 저장기간이 길어지면서 보다 깊게 휴면에 이르는 경향이었다(Fig. 2B). 한편 발아속도에 있어서도 갯줄풀은 휴면타파 저장기간이 길어질수록 보다 빨라져, 100일 저온 해수침지한 경우 발아종료일이 7일로 단축되었지만 영국갯끈풀의 발아속도 단축은 매우 완만한 경향을 나타냈다(Fig. 3). 휴면타파조건의 경우도 갯줄풀은 해수 및 담수간의 차이가 거의 없었지만 영국갯끈풀의 경우에는 담수에서의 휴면타파가 해수에서 보다 불량한 편이었다(Fig. 2). 그러나 가장 우수한 종자휴면타파는 두 종 공히 해수에서 일어났고, 대체로 3개월 이상이면 충분히 타파되었다(Fig. 2). 이들의 결과를 볼 때, 종자로부터의 번식은 영국갯끈풀보다 갯줄풀이 상대적으로 보다 양호할 것으로 여겨지기 때문에 이에 대한 고려가 필요할 것이다. 그런데 갯줄풀 종자의 휴면 여부에 대해서는 서로 상반된 결과들이 보고되고 있다. 갯줄풀 종자는 수집 후 바로 10-29%의 발아를 나타내었고 (Elsey-Quirk et al., 2009), 휴면은 관찰되지 않았으며(Mooring et al., 1971), Li et al. (2010)는 갯줄풀 종자가 휴면의 징조없이 증류수 및 pH 6.63-9.95의 저염도(≤200 mM)에서도 발아율에 큰 차이 없이 70% 이상 매우 양호하였다고 하였다. 그러나 Biber and Caldwell (2008)의 보고에서는 갯줄풀 종자가 휴면이 있으며 4℃ 암조건의 저온습윤 저장(stratification) 처리시 1개월째부터 발아되기 시작하여 4개월째 35% 정도로서 가장 높았다가 이후 감소하는 경향을 보였다고 하였다. 이러한 경향은 타 연구자의 결과(Yuan and Shi, 2009)에서도 관찰되었다. 그리고 Marks and Truscott (1985)의 연구에 의하면 영국갯끈풀 종자도 휴면이 있어서 5℃ 저온 습윤조건의 60일 저장에 의해 발아가 촉진되었다고 하였다. 본 연구에서도 S. alterniflora 종자가 강한 휴면성이 있음을 확인하였는 바, 연구자들간의 서로 상반된 연구결과의 원인에 대해서는 추후 면밀한 비교조사가 이루어져야 할 것이다.
한편 발생적인 측면에서 cordgrass는 몇가지 유리한 점을 가지고 있는 것 같다. 즉 발아에 광이 필요치 않으며(Fig. 4), 비교적 고온조건(낮 35℃/밤 25℃, 12 h 광주기)에서도 발아가 가능하고(Fig. 5), 염도가 높을수록 발아율이 저감되기는 하지만 6% 미만의 높은 염도조건에서도 쉽게 사멸되지 않고 발아력을 보유한다는 것이다(Table 2). 또한 양분결핍, 온도, 얕은 침수 등과 같은 불량환경조건에서의 생존력이 상대적으로 우수한 것 같다. 이는 휴면타파된 이후엔 1년내내 육상 또는 해안에서 광범위하게 발생할 수 있는 잠재력이 있다는 것을 의미한다. 이와 반면에 발생 측면에서의 단점도 있는 것 같다.
즉, 종자는 2차 휴면이 없어 (Table 1) 갑자기 건조 조건에 접하면 쉽게 생명력을 잃는 것 같으며(Goodman et al., 1969; Mooring et al., 1971; Plyler and Carrick, 1993; Probert and Longley, 1989), 저산소함량(2.5와 5%) 조건에서 종자 발아율이 감소되었고(Wijte and Gallagher (1996), 종자로부터의 유묘 출현율은 4 cm 수심부터 현저히 감소하였고 생존율도 5% 미만을 나타냈으며(Abbas et al., 2012), 성숙 식물체도 장기간의 침수(flooding)에 의해 용이하게 고사된다고 한다(Smith and Lee, 2015; Yuan et al., 2011). 이외에 다른 한가지의 특성으로서는 영국갯끈풀의 초기 생장속도가 Lolium perenne 보다 매우 느렸고(Dunn et al., 1987) 본 연구에서도 갯줄풀 발아 이후 유묘의 생장이 느린 것으로 관찰되었는 바(Fig. 8) 타 식물과의 경합력이 있는 조건에서는 이들이 상대적으로 약하게 생장할 것 같다. 이는 타 식물이 잘 자라지 못하는 해안가에서 cordgrass가 손쉽게 우점력을 확보하는 것도 이러한 특성때문으로 여겨진다. 그렇지만 일단 정착된 식물은 강한 군락을 형성하면서 불량환경속에서도 잘 적응하면서 근경 및 종자로 많은 량이 번식되기 때문에 무엇보다 초기 관리에 유념해야 할 것이다.

