서 언
국내에서 억새(Miscanthus spp.)는 다년생 화본과 C4 식물로서 과수원과 비농경지에서 비선택성 제초제인 glyphosate로도 방제가 어려운 잡초로 인식되어왔다. 그러나 2000년대 들어 억새는 경관생태용으로 각광을 받기 시작하였으며, 2010년대에는 바이오에너지 작물로서 개발을 위한 다양한 연구가 진행되어왔다. 바이오에너지 작물로서 관심을 끌고 있는 주요 억새종은 대체로 동아시아 지역에 분포하며 국내에는 3개 종(M. sinensis,M. sacchariflorus 및 M. floridulus)이 전국적으로 자생하는 것으로 보고되었다(Chung and Kim, 2012; Hodkinson et al., 2002a). 억새는 높은 바이오매스 생산성을 갖고 있으면서도 환경적응성이 뛰어나 해외에서는 이미 바이오에너지 작물로 상업적 재배가 이루어지고 있으며, 형태적으로도 아름다워 오래 전부터 조경용으로 재배되고 있다(Clifton-brown et al., 2011; Stewart et al., 2009). 억새는 생초 생산량이 많아 사료용 작물로서 개발 가능성이 있으며, 억새의 빠른 생장 및 피복능력은 생태복원용으로의 활용 가능성도 높은 것으로 판단된다.
억새의 재배에 있어 장애요인은 번식이 주로 근경에 의존하고 있으며, 종자를 통한 번식 및 재식 방법이 충분히 확립되어 있지 않다는 것이다. 근경을 통한 번식 및 재식은 많은 시간과 노동이 투입되므로 관련 비용이 많이 소요되며(Lee, 2010), 이는 특히 상업적 재배를 위한 대량번식 시 억새의 작물로서의 경쟁력을 약화시킨다. 이를 극복하고자 조직의 기내배양을 통한 배양 묘, 줄기 마디, 플러그 묘 등을 생산하는 다양한 번식 방법이 연구되어 왔으나 유의한 성과를 거두지 못하였다(Atkinson, 2009; Hwang et al., 2018; Xue et al., 2015). Xue et al. (2015)이 보고한 바에 따르면 억새의 근경, 줄기 및 종자 기반의 번식 방법 중 종자를 통한 번식이 대규모 포장 조성 시 가장 경제성이 높은 것으로 분석되었다. 그러나 억새 종자는 발아가 균일하지 않고 발아율이 낮아 입묘가 어려워 상용화하기에 한계가 있다(Christian, 2012; Christian et al., 2005). 억새가 자생하지 않는 유럽국가의 경우 1930년대 일본에서 건너간 2배체 참억새(2n=2x=38)와 4배체 물억새(2n=4x=76)의 자연교잡종인 3배체 M. x giganteus (2n=3x=57)가 바이오매스 생산용으로 재배되고 있으나(Hodkinson et al., 2002b; Linde-Laursen, 1993), 불임으로 종자번식이 불가하여 동아시아에서 수집한 2배체 참억새의 교잡을 통해 종자번식이 가능한 참억새 품종을 개발하고 있다. 우리나라는 억새의 자생지로서 다양한 유전자원, 특히 종자생산이 가능한 2배체 참억새와 2배체 및 4배체 물억새 유전자원이 자생하고 있어(Lim et al., 2014; Yook et al., 2014), 종자번식이 가능한 억새품종 개발이 용이하다. 그러나 이들 억새의 종자발아특성에 대한 연구가 충분히 수행되지 않아 종자를 기반으로 한 대량증식 방법이 확립되어 있지 않고 적합한 발아조건 등에 대한 정보가 부족한 실정이다. 따라서 종자번식하는 억새 품종의 개발 및 상용화를 위해 억새 종자의 발아를 제고하기 위한 방법의 개발이 시급히 요구된다.
종자의 발아를 제한하는 요인은 배 미숙, 종피의 불투성, 휴면물질의 존재 등으로 매우 다양하며(Baskin and Baskin, 1998), 억새 종자의 발아가 불량한 원인은 명확히 밝혀진 바 없기에 억새 종자의 발아를 향상시키는 발아 조건 확립을 위해 일반적으로 종자처리에 사용되는 지베렐린(GA3), KNO3 및 종자소독제 등 외부 화합물을 우선적으로 적용하여 그 효과를 평가할 필요가 있다. 따라서 본 연구는 GA3, KNO3등의 발아촉진제와 NaClO, prochloraz 등의 종자소독제 처리에 따른 억새 종자의 발아특성을 평가하여 외부 화합물의 발아 촉진 효과를 확인하고 최적 종자처리 방법을 도출하고자 수행되었다.
재료 및 방법
2009년 10월 억새 유전자원포장에 재식후 2년이 된 물억새 2종(SNU-M-004, 040)과 참억새 1종(SNU-M-044)으로부터 건실한 종자를 수확하여 실내에서 자연 건조한 후 실험에 사용하였다. 사전 예비 발아실험에서 종자의 영이 존재할 경우 최종 발아율과 속도가 매우 저해되었으므로 전처리 과정에서 외영과 내영을 제거하였으며, 미숙립과 피해립도 제거하였다. 화합물에 따른 억새 종자의 발아특성 평가를 위해 페트리디쉬에 여과지를 깔고 각각의 정상립 50립씩을 파종하였다. 억새 종자에 대한 외부 화합물 처리는 다음과 같이 수행되었다. GA3와 KNO3는 물억새(SNU-M-040) 및 참억새(SNU-M-044)를 대상으로 처리 농도별로 조제한 용액을 페트리디쉬에 주입한 후 종자를 치상하고 증발한 만큼의 수분을 수시로 보충하였다. GA3의 처리 농도는 0, 2, 10, 50, 250 mg/L, KNO3는 0, 1, 10, 100, 1000 mM이었다. 종자소독제로 사용되는 NaClO 및 prochloraz는 처리 농도별 용액에 물억새(SNU-M-004) 종자를 1시간 동안 침지시킨 후 증류수에 3-4회 세척 후 페트리디쉬에 파종한 후 종자가 과습하지 않을 정도로 적정량의 수분을 수시로 공급하였다. NaClO의 처리 농도는 0, 0.3, 0.6, 1.2, 2.4%이었고, prochloraz는 0, 0.01, 0.1, 1, 10%이었다. 각 처리구는 3반복으로 완전임의배치하여 30℃ (14 h/ 주간)-20℃ (10 h/ 야간)의 온도 조건을 유지하였다. 파종 후 3일째까지 매 8시간 간격, 4일째부터 10일째까지 24시간 간격으로 발아 개체 수를 조사하였으며, 유근이 2 mm 이상 나타난 종자를 발아된 것으로 간주하였다.
종자의 파종 후 시간(t)에 따른 누적 발아율(y)은 비선형 회귀분석을 통해 식물의 발아, 출아 및 생장율 분석에 효과적인 Gompertz 모델(식 1)에 적합시켜(Gompertz, 1825; Kim et al, 2006; Steppunn et al., 1998) 최대누적발아율(C), 누적발아곡선의 기울기(B), 최대누적발아율(C)의 50%에 도달하는데 소요되는 기간(M)을 구하였다. 아울러 Gompertz 모델과 추정된 모수(C, B, M)를 이용하여 90% 발아 도달에 소요되는 기간(T90)을 계산하였다. 모든 통계분석은 Genstat (Genstat 5 Committee, 1997)을 이용하여 실시되었다.
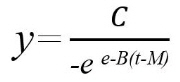 (1)
(1)
결과 및 고찰
억새 종자발아에 미치는 GA3 효과
GA3 처리가 억새 종자 발아에 미치는 영향을 평가하고자 시간에 따른 누적발아율을 Gompertz 모델에 적합시키는 비선형회귀분석을 수행한 결과, 물억새와 참억새 두 종 모두 무처리 대비 모든 처리 농도에서 발아 속도가 향상되었다(Table 1; Fig. 1). 물억새의 경우 무처리(24시간)를 제외한 모든 처리구에서 최초 발아에 소요된 시간이 16시간으로 단축되었으며, Gompertz 모델로 예측한 최대 발아율의 50%에 도달하는 시간(M) 및 초기 발아속도(B)는 무처리에서 26.6시간 및 0.151었으나 2, 10, 50 및 250 mg/L GA3 처리 시 각각 19.6, 18.1, 16.5, 17.7시간과 0.250, 0.221, 0.369, 0.208로 추정되어 GA3 처리에 의해 발아세가 높아지는 효과를 확인하였다. 참억새의 경우 최초 발아 개체가 조사된 시간은 무처리와 2 mg/L GA3 조건에서 24시간이었고 10 mg/L GA3 이상의 농도를 처리했을 때는 파종 후 16시간 후에 발아한 개체를 볼 수 있었다. 최대 발아율의 50%에 도달하는 시간(M) 및 초기 발아속도(B)는 무처리에서 27.6시간 및 0.247이었으며 2, 10, 50 및 250 mg/L GA3 처리 시 각각 23.9, 22.1, 22.4, 19.5시간과 0.331, 0.412, 0.271 및 0.220으로 추정되어 GA3 처리에 의해 발아세가 증가하는 것으로 나타났다.
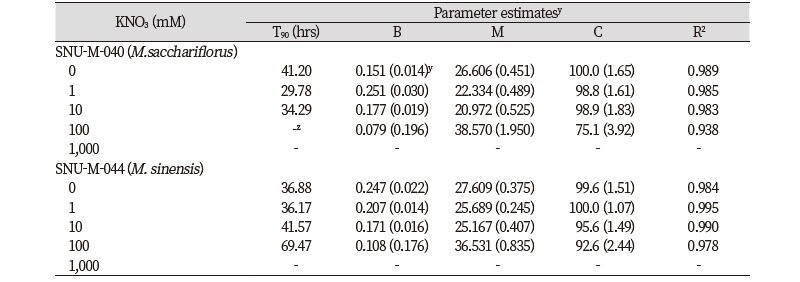
이는 참억새의 발아휴면특성에 관한 기존 연구에서 GA3 처리에 의해 발아 속도가 증가한다는 결과와도 일치하였다(Aso, 1976; Christian, 2012, Christian et al., 2014). 물억새를 이용한 발아특성실험은 매우 제한적이었으나 본 연구 결과를 통해 참억새와 유사하게 GA3에 의해 발아가 촉진되는 것을 밝힐 수 있었다. 특히 최종 발아율의 90%에 도달하는 시간(T90)이 물억새는 무처리 대비 최대 45% 감소(50 mg/L GA3)하여 25% (10 mg/L GA3) 감소한 참억새보다 더 큰 발아 촉진 효과가 나타났다. 이상의 결과를 통해 GA3 처리 시 물억새의 최적 발아 조건 50 mg/L, 참억새는 10 mg/L 농도가 최적 발아 조건인 것을 확인할 수 있었다.
억새 종자발아에 미치는 KNO3효과
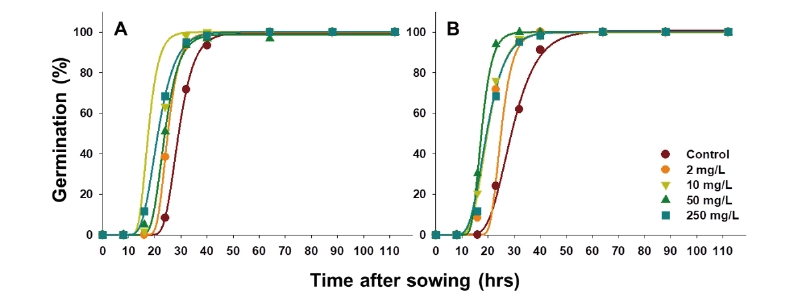
물억새와 참억새 종자에 KNO3를 처리하여 발아에 미치는 영향을 평가한 결과에 따르면 저농도 처리는 초기 발아 속도를 증가시키나 그 효과가 미미하였고, 100 mM 이상에서는 발아율 및 발아 속도가 모두 저해되어 1,000 mM에서는 발아한 개체가 없었다(Table 2; Fig. 2). 물억새의 경우 최초 발아 개체가 조사된 시간은 무처리는 24시간, 1, 10 mM 조건에서 16시간으로 짧아졌다가 100 mM 조건에서 다시 24시간으로 증가했다. Gompertz 모델로 예측한 최대 발아율의 50%에 도달하는 시간(M) 및 초기 발아속도(B)는 무처리에서 26.6시간 및 0.151이었으며 1, 10 및 100 mM KNO3처리 시 각각 22.3, 21.0, 38.6시간과 0.251, 0.177, 0.079로 농도 증가에 따라 발아세가 떨어지는 경향을 보였다. 참억새의 경우 KNO3처리에 따른 유의미한 발아 촉진 효과를 찾을 수 없었다.
이상의 결과와 T90 추정값을 고려할 때 무처리 대조군 대비 KNO3처리에 의한 발아 촉진 효과는 저농도 처리 시 물억새에서만 미미하게 나타났다. KNO3는 일반적으로 종자의 수분 상태를 조절하는 기능을 하는 것으로 알려져 있으나 정확한 발아 촉진 기작은 밝혀져 있지 않다. 따라서 원인을 특정하긴 어려우나 0.2% KNO3를 참억새 종자에 처리했을 때 발아율이 대조군에 비해 50% 이상 감소한 기존 연구 결과(Cho et al., 2012)를 고려했을 때 KNO3는 억새 종자 처리 방법으로 적합하지 않다고 판단된다.
억새 종자발아에 미치는 종자소독제의 효과
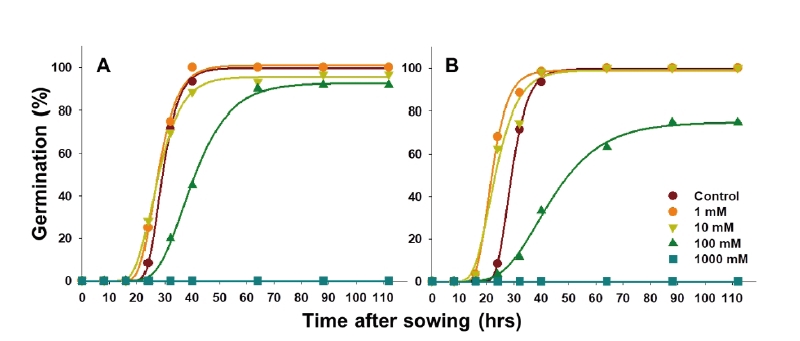
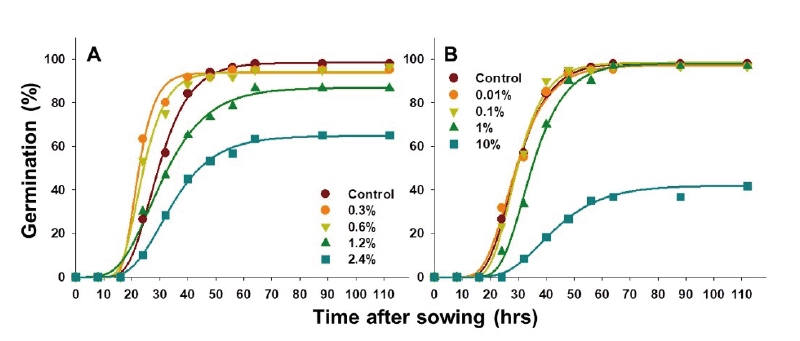
NaClO를 물억새 종자에 전처리하여 발아 특성을 평가한 결과, 0.3%와 0.6% 처리에서 발아 촉진 효과가 나타났으나 농도가 높을수록 발아율 및 발아 속도가 감소하였다(Table 3; Fig. 3A). 모든 NaClO 처리구의 최초 발아 소요 시간은 무처리 조건과 동일하게 24시간으로 관측되었다. Gompertz 모델로 예측한 최대 발아율의 50%에 도달하는 시간(M) 및 초기 발아속도(B)는 무처리에서 26.6시간 및 0.129였으며 0.3 및 0.6% 처리 시 각각 20.7, 21.8시간과 0.247, 0.179로 저농도 조건에서는 발아세가 다소 증가하는 것으로 나타났다. 그러나 NaClO 농도가 높아질수록 종피가 녹는 것이 확인되었고 1.20% 이상의 농도에서는 종피뿐만 아니라 종자도 일부 피해를 입으면서 발아가 급격히 저해되어 최대 누적 발아율이 64.7%까지 감소하였다. 따라서 강염기인 NaClO에 의한 종피의 피해를 최소화하면서 발아를 촉진하기 위한 최적 처리 농도는 0.3%로 평가되었다.
한편 종자소독제로 이용되는 Prochloraz의 경우 모든 처리 농도에서 발아가 촉진된다는 유의적인 차이를 발견 할 수 없었으며, 오히려 1%에서는 초기 발아 속도가 감소하고 10% 처리 시에는 최종 발아율마저 50% 이상 감소하여 물억새 종자의 발아 증대를 위한 처리로 부적합하였다(Table 3; Fig. 3B).
억새 종자발아 최적 조건
억새 종자의 휴면 요인은 실험적으로 명확히 밝혀지지 않았으나 Baskin and Baskin (1998)에 따르면 일반적으로 화본과 초본류 종자의 경우 물리적 휴면 및 형태적 휴면이 없고 대체로 생리적 요인에 의한 휴면만 존재하며, 대부분은 건조 저장, 외영 및 내영 등을 물리적으로 제거 또는 GA3 처리를 통해 발아가 촉진된다. 본 연구에서는 실험 수행 이전까지 종자를 자연 건조 상태로 보관하고, 영을 제거한 억새 종자를 사용하였으므로 Baskin and Baskin (1998)이 제시한 화본과 초본류 종자의 휴면 타파 방법 중 두 가지가 선행된 상태에서 실험을 수행하였다. 이에 따라 무처리 대조군의 최종 누적 발아율이 대체로 100%에 도달하여 외부 화합물의 처리 효과가 과소평가되었을 가능성이 있다. 실제 대량번식을 위한 육묘 시에는 영을 제거하기가 어려우므로 본 연구에서 제안된 억새 종자 처리 방법이 영을 제거하지 않은 조건에서 어떤 양상으로 나타날지 추후 연구가 이루어져야 할 것이다. 한편 억새 종자로부터 플러그묘를 육묘한 선행연구에 따르면 발아 속도가 빠른 품종이 뿌리가 발달하여 토양과 잘 응집해서 이식 가능한 플러그를 형성하는 시기에 빠르게 도달하였으므로(Hwang et al., 2018), 화합물 처리를 통해 억새 종자의 초기 발아 속도를 증대시키는 것이 억새 육묘 시 비용과 시간을 절감하는 효과를 가져올 것으로 예상할 수 있다. 따라서 억새 종자의 휴면을 타파 및 발아율 향상을 위해 2종의 발아촉진제와 2종의 종자소독제를 평가한 결과 물억새 종자는 저농도의 KNO3및 NaClO 처리 시에도 발아가 다소 촉진되었으며, GA3 처리가 참억새 및 물억새의 발아 촉진에 가장 효과적이었다. 본 연구의 결과를 종합적으로 고려하였을 때 물억새의 종자 발아 향상을 위한 최적 처리 조건은 50 mg/L GA3, 참억새는 10 mg/L GA3 처리가 가장 우수한 처리 방법으로 평가되었다.
억새는 환경에 대한 적응력이 높고 생장 속도가 빠른 특성으로 인해 작물 재배가 어려운 한계 농지에서 바이오매스의 생산 또는 생태복원에 적합하나, 근경을 통한 영양 번식의 한계로 인해 상용화에 어려움이 있다. 따라서 본 연구에서 확립한 GA3 기반의 종자 처리방법을 통해 종자를 통한 억새의 번식 효율을 높인다면 억새의 대규모 재배 생산의 효율성과 경제성을 제고할 수 있을 것으로 기대된다. 또한 억새와 유사한 화본과 식물의 종자 휴면성 타파 및 발아율 향상 및 종자기반 대량증식에도 응용 적응할 수 있을 것으로 기대된다.

