서론
1992년 생물다양성 협약(CBD: Convention on Biological Diversity)이 체결되며 생물자원의 국가 소유 권리가 인정되었다(Yu, 2006). 이후 국내외에서 생물자원에 대한 주권 확보와 생물다양성 보존을 위한 노력이 증가하고 있다(Yu, 2006; Lee et al. 2013). 침입 외래생물은 생물다양성 감소에 큰 영향을 미치는 요인 중 하나로 기존 생태계에 유입될 경우, 자생식물의 종 다양성을 감소시키고, 생태계 서비스에 부정적인 영향을 미친다(Kil and Kim, 2015). 또한 농업, 임업, 산업 분야에서 상당한 경제적, 사회적 피해를 일으킨다(Rebek et al. 2006; Park and Lee, 2018; Kim and Koo, 2021).
나고야 의정서(Nagoya Protocol)와 쿤밍-몬트리올 글로벌 생물다양성 프레임워크(K-M GBF: Kunming-Montreal Global Biodiversity Framework)에서 ’침입 외래생물의 관리 및 제거’에 대한 권고와 같이 침입 외래생물의 효율적인 관리를 위한 국제적 규범화가 활발히 진행되고 있다(Kim, 2023). 이에 따라 각 국가는 침입 외래생물을 목록화하고, 이를 관리하기 위해서 연구 수행과 관리 방안을 구축하고 있다(Park and Lee, 2018).
국내의 경우 국가생물다양성 전략을 수립하여 침입 외래생물의 조사와 방제 강화를 권고하고 있다. 또한 「생물 다양성 보전 및 이용에 관한 법률」(이하 ʻ생물다양성법’) 시행을 통해 생태계에 대한 교란이 크거나 우려가 있는 외래생물을 생태계교란 생물, 유입주의생물, 생태계위해우려생물로 지정하여 관리하고 있다(Kim and Koo, 2021). 그리고 생물다양성법 제24조와 제27조에 의거하여 매년 생태계교란 생물 퇴치사업과 방제 기술개발 사업을 추진하고 있다(Park and Lee, 2018; Kil and Kim, 2015).
하지만 침입 외래생물의 지속적인 규제와 관리에도 불구하고 관련 생물의 생태적 지식과 모니터링 자료, 전문 인력의 부족으로 종별 또는 발생지역별 맞춤형 방제체계 구축이 어려워 제거 기술개발과 퇴치의 실효성이 매우 낮은 실정이라고 보고되었다(Park et al., 2016). 그리고 침입 외래생물의 생리, 생태적 변화는 종별로 다르게 나타나기 때문에 모든 침입 외래생물에 일반화된 관리 전략을 사용할 수 없다(Park et al., 2016).
따라서 침입 외래종의 방제는 생활사 연구를 통한 방제 시기 설정, 생리·생태적 특성에 관한 연구를 통한 종별 관리 전략이 수립되어야 한다(Kim and Koo, 2021). 또한 종의 생리, 생태적 특성과 지역에서의 생태적 특성을 고려하여 이루어져야 한다(Park et al., 2015).
마늘냉이(Alliaria petiolata)는 유라시아 원산지의 월년생 십자화과 식물이다. 현재 국내에는 강원도 삼척, 경기도 수원, 인천 중구, 충남 당진에 분포하고 있다(Kim et al., 2021, Yun et al., 2022). 마늘냉이는 생태계교란 생물 중 하나로 2020년에 지정되었다. 미국에선 유해 잡초로 등록되어 있고, 캐나다에선 생태적 피해가 커 유해식물로 지정하여 관리하고 있다(Kim et al., 2021). 마늘냉이는 한 개체 당 15,000개의 종자를 생산하며 번식력이 높고 확산이 빠르다. 그리고 대규모 군락을 형성하여 숲 하층을 우점해 기존 식생의 생물다양성을 감소시킨다(Anderson et al., 1996; Kim et al., 2021, Yun et al., 2022).
마늘냉이는 비교적 최근에 국내 생태계에 침입하여 분포지가 제한적이다(Yun et al., 2022). 생태계교란 생물의 경우 적응, 생장과 번식력이 기존 자생식물보다 빠르기 때문에 방치할 경우 우점화를 통해 여러 생태적 문제를 일으킨다. 따라서 발생 초기에 신속히 제거하는 것이 중요하다(Kim, 2016).
본 연구는 마늘냉이의 대규모 확산 전 제거 및 관리 방안 수립을 위해서 필요한 생육특성을 다양한 차광조건에서 생장 및 생리적 반응 차이를 확인하여 생태적 방제 방법 개발에 필요한 기초자료 제공과 효율적인 물리적 방제 방법을 모색하고자 수행하였다.
연구재료 및 방법
1. 시험재료 및 차광 처리
실험용 재료인 마늘냉이(Alliaria petiolata)는 2023년 3월에 마늘냉이 서식지인 광교 호수공원(경기도 수원시 영통구 하동 490) 부근에서 채집하였다. 채집한 마늘냉이는 크기별로 분류하여 길이가 15~21 cm인 개체를 사용하였다. 선별된 개체는 60×40×30 cm 포트에 20 cm 간격으로 6개체씩 재식하였으며, 차광 처리구에서 처리 방법별로 4 반복씩 288개체가 사용되었다. 재식된 마늘냉이는 온실에서 3주간 활착시킨 후 차광 처리를 진행하였다. 시험에 사용된 토양은 시판 준중량 상토 20 L 와 가축분퇴비 1 L를 혼합 후 사용하였다.
본 시험은 한경국립대학교(경기도 안성시 중앙로 327) 구내에 조성된 마늘냉이 차광 시험구에서 실시하였다. 차광망은 시중에서 판매되고 있는 흑색 차광망을 사용하였으며, 직사광을 그대로 받는 대조구와 35% 차광막 설치구, 75% 차광막 설치구 등 3가지 조건으로 진행하였다. 차광 시험구 별 환경조건을 알아보기 위하여 대기 온습도 센서를 연결한 데이터 로거(Model E1650, Model E1450, Echo Korea)를 시험구 중앙의 1.5 m 높이에 각각 설치하여 대기 온습도를 매일 1시간 간격으로 8주 동안 측정하였다. 광합성광양자량(Photosynthetic Photon Flux Density: PPFD)은 휴대용 광합성 측정 장치(OHSP350P Spectral Irradiance Colorimeter)를 이용하여 각 차광 시험구에서 3월 30일부터 6월 1일까지 5회 네 지점을 측정하여 평균값을 사용하였다.
2. 차광조건에 따른 생육특성
생장 특성 중 초장(cm), 엽장(cm), 엽폭(cm), 엽수(ea), 엽록소 형광의 조사는 차광 처리 후 2023년 4월 1일부터 6월 1일까지 10일 간격으로 진행하였다. 각 처리구에서 4 반복으로 조사하였다. 생체중(g)과 건물중(g) 측정은 재식 8주 후 진행되었다. 생체중(g)을 측정한 후 80℃에서 48시간 건조 후 건물중(g)을 측정하였다. 엽면적지수(cm2)는 재식 4주 후 식물체의 중간 부위에서 완전히 전개된 잎 중 4엽씩 채집한 뒤 ʻimage j’ 프로그램을 활용하여 측정하였다. 비엽면적(SLA, Specific Leaf Area)은 엽면적을 측정한 잎을 건조 후 생체량을 측정하여 엽 생체량 당 엽면적의 비로 산출하였다(Hwang et al., 2003). 엽록소 형광 반응(Origin Jump Intermediate Peak; OJIP) 측정은 식물체의 중간 위치에 있는 완전히 전개된 잎을 대상으로 Fluorepen FP-100 (PS I, Czech)을 사용하여 측정하였다. 이때, 엽록소 형광 반응 측정 전 식물체의 잎맥을 제외한 중간 부위를 leaf clip을 활용하여 30분간 암 처리 후 측정하였다.
엽록소 함량 측정은 차광 처리 4주 후 진행하였으며, 각 처리구 별로 3장의 잎을 채집하여 0.5 cm2 정사각형으로 절단하였다. 색소 추출을 위해 절단한 잎 0.5 g과 95% ethanol 5 ml를 50 ml falcon tube에 넣은 후 4℃에서 24시간 동안 암처리 하였다. 원심분리기(General Purpose Centrifuge VS-4000N, Vsion Science, Korea)를 이용하여 3,000 rpm에서 10분간 원심분리를 하고, 상등액 1 ml를 blank와 함께 UV-VIS Spectometer(T6 UV-VIS spectrometer, PERSEE, USA)를 사용하여 649, 664, 750 nm 파장에서 흡광도를 측정하였다. 측정된 흡광도를 공식에 따라서 엽록소 함량을 산출하였다(Winterman and de Mots, 1965).
- Chlorophyll a=[(13.7×(A665-A750)-5.76×(A496-A750))×C×V×D]/(W×1000)
- Chlorophyll b=[(25.8×(A665-A750)-7.6×(A496-A750))×C×V×D]/(W×1000)
Pigment Equation(C=measuring volume (mL), V=extraction volume(mL), D=sample dilution factor, W=fresh weight of sample).
3. 차광조건에 따른 제거 효과
시험은 세 가지 차광조건(0%, 35%, 75%)에서 진행되었다. 마늘냉이 제거 방법은 4월에 지상부 제거, 5월에 지상부 제거, 4월에 지하부 절단 및 대조구 등 4가지 방법으로 진행하였다. 95% ethanol로 소독된 가위를 이용하여 마늘냉이의 지상부, 지하부를 절단하였다. 지상부와 지하부의 경계점은 원뿌리와 줄기의 경계인 뿌리목을 기준으로 처리를 진행하였다. 처리 8주 후 고사율과 종자 수량을 조사하여 처리에 따른 제거 효율을 비교하였다.
고사율은 각 차광 시험구에서 처리 방법별로 4 반복으로 조사하였다. 고사 여부의 판단은 절단된 마늘냉이의 뿌리에서 줄기 및 잎의 재발생 여부에 따라 판정하였다. 고사율은 ʻ(고사한 마늘냉이 수/식재 마늘냉이 수)×100(%)’로 변환하여 사용하였다.
종자 수량은 각 차광 시험구의 처리 방법별 생산된 마늘냉이 종자를 채집하여 조사하였다. 종자 수량은 전체 무게와 천립중을 측정하여 ʻ(전체 무게/천립중)×1,000’으로 변환하여 전체 예상립수를 추정하였다(Gang et al., 2019).
4. 통계분석
통계 처리는 SAS(SAS Institute Inc. Cary, USA)의 PROC ANOVA를 활용하여 분산분석을 실시하였고, 다중범위검정법(Ducan’s multiple range test, DMRT)을 이용하여 평균 간의 통계적 유의성을 검증하였다.
연구결과 및 고찰
1. 차광조건별 환경인자
실험기간 동안 각 처리구 내부의 월별 평균 대기온도(℃), 대기습도(%)를 조사한 결과, 대조구의 경우 4월 14.37℃, 58.26%, 5월 19.86℃, 55.84%, 6월 25.18℃, 54.52%로 조사되었다. 처리구 중 35% 차광처리 시험구는 4월 13.64℃, 57.28%, 5월 19.31℃, 55.00%, 6월 23.65℃, 53.38%로 조사되었으며, 75% 차광처리 시험구의 경우 4월 12.53℃, 56.89%, 5월 19.56℃, 54.70%, 6월 23.64℃, 53.27%로 측정되었다(Fig. 1).
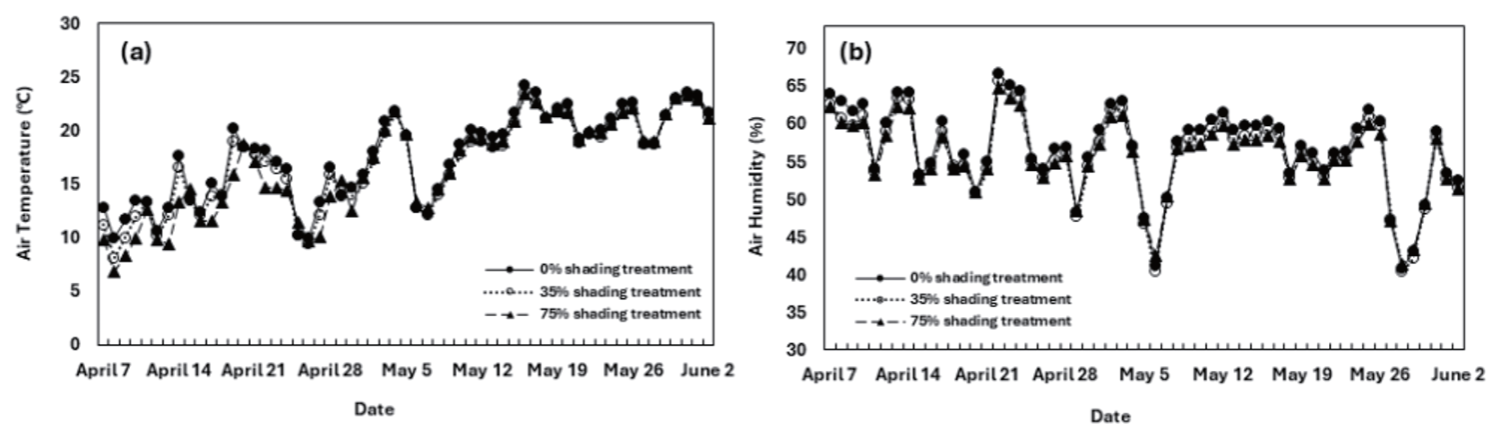
광합성광량자속밀도(Photosynthetic Photon Flux Density: PPFD)는 광합성에 직접적인 영향을 미치는 광량자속이 초당 평방미터에 떨어지는 양을 말한다. 처리구 내부의 평균 PPFD는 대조구 779.18μmol·m-2·s-1, 35% 차광처리 시험구는 353.00 μmol·m-2·s-1, 75% 차광처리 시험구는 231.00 μmol·m-2·s-1로 측정되었다(Table 1). 대조구에 비하여 35% 차광처리 시험구는 1.5배 더 낮았고, 75% 차광처리 시험구는 대조구와 비교하였을 때 약 3.5배 더 낮은 값을 보였다.
Table 1
List of predicted transcription factors interacting APX genes in four species of zoysiagrass.

2. 차광처리에 따른 생육특성
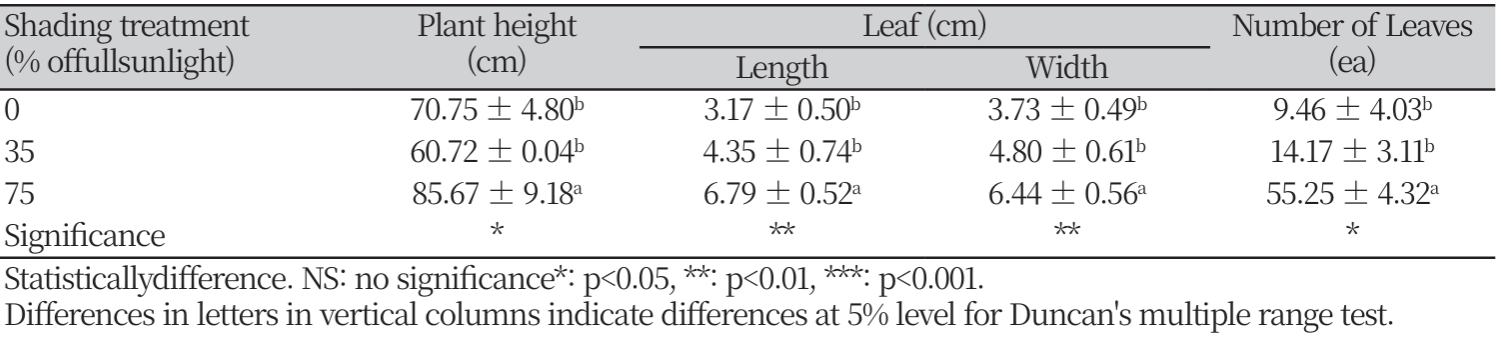
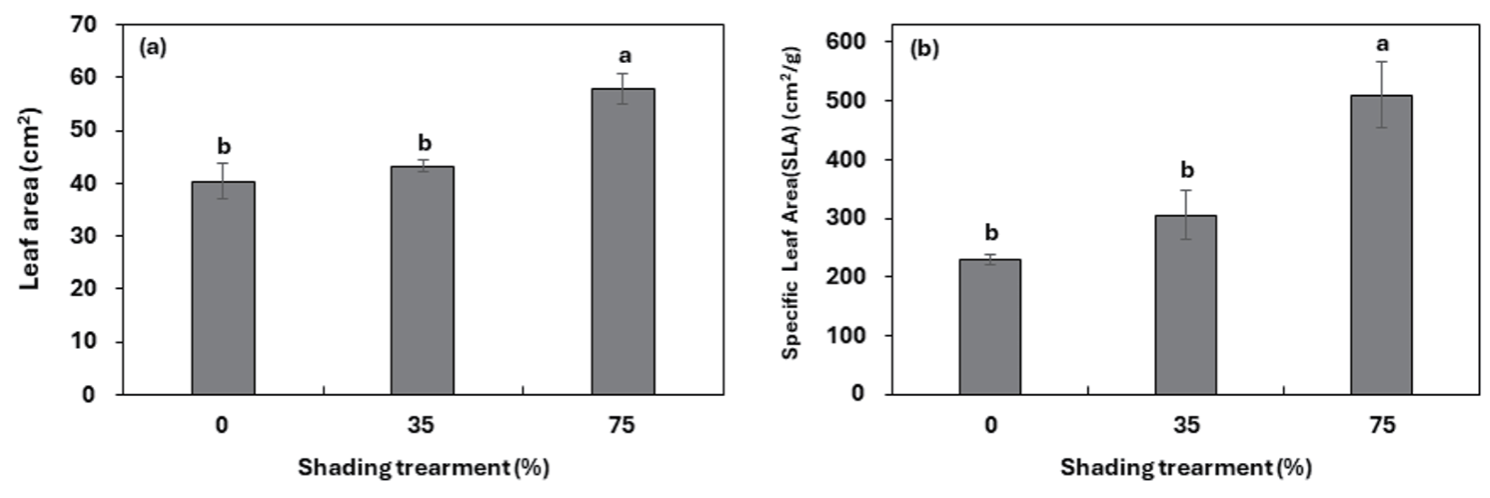
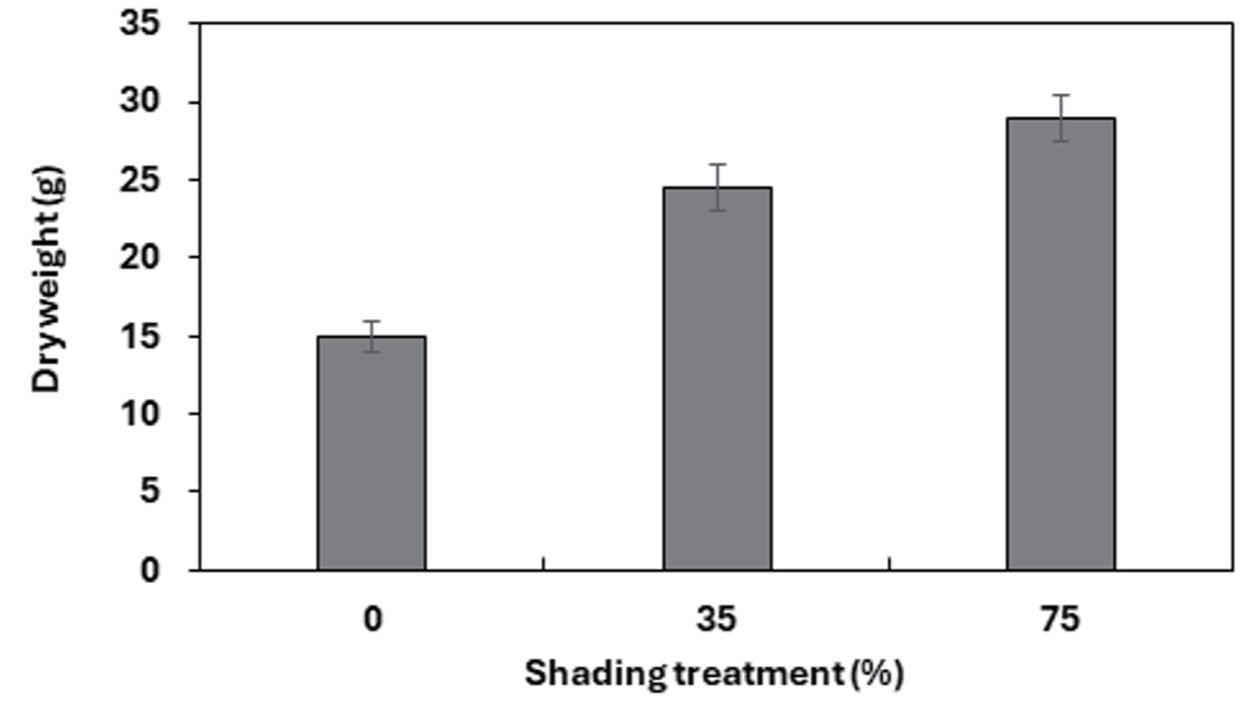
차광조건에 대해서 모든 인자들은 고도의 유의성을 보였다. 차광조건별 모든 생장 특성에서 대조구는 75% 차광조건과 유의한 차를 보였지만, 대조구와 35% 차광조건 사이의 통계적 유의차를 나타내지 않았다. 마늘냉이는 전반적으로 차광률이 높을수록 초장, 엽장, 엽폭, 엽수와 엽면적, 비엽면적(SLA: Specific leaf area)이 증가하였는데(Table. 2, Fig. 2), 이는 내음성 수종들은 음지에서 제한된 빛을 이용한 광합성 수행 능력을 갖기 위하여 차광조건에서 형태학적으로 양지식물과는 다른 특성 차이를 보이는 것으로 판단된다(Je et al., 2006; Lee et al., 2020). 본 결과는 내음성 식물의 경우 광도가 증가할수록 생장률이 낮아진다는 Kobe et al (1995)의 연구와 같은 결과를 보였다. 차광 처리 8주 후 대조구 대비 75% 차광조건에서 초장은 약 1.2배, 엽장과 엽폭은 2배, 엽수는 약 6배 더 높은 값을 보였다. 본 결과는 낮은 광도에서 자란 잎은 면적을 넓게 하여 수광량과 빛의 흡수를 높이기 위한 것으로 판단된다(Adams et al., 1990; Evans, 1994). 엽수의 경우 내음성이 강한 식물의 경우 차광률이 높아질수록 엽수가 증가한다는 Yoo and Kim(1997)의 연구와 Choi et al (2012)의 실험과 같은 결과를 보였다. 차광처리 4주 후 엽면적과 비엽면적(SLA)은 차광률이 높아질수록 증가하였으며, 대조구와 비교하였을 때 75% 차광조건에서 약 1.5배, 약 2배 더 높게 나타났다. 이와 같은 결과는 음지식물인 곰취의 경우 차광률이 높아질수록 비엽면적이 증가했다는 Song et al (2014)의 실험과 같은 결과를 보였다. 내음성이 강한 식물은 빛을 받는 엽면적을 최대화하기 위해서 양지식물보다 잎을 얇고 넓게 전개시켜 더 높은 비엽면적(SLA)을 갖는 것으로 판단된다(Hong and Yoon, 2000; Farnworth and Ellison, 1996; Poole et al., 1996). 이는 잎의 두께를 줄이는 대신 면적을 넓게하여 광합성을 많이 하도록 함으로써 음지에서 적응력을 높이기 위한 방법으로 사료된다Yoo and Kim(1997). 마늘냉이의 건물중은 차광조건이 증가할수록 건물중이 15.02g, 24.48g, 29.00g으로 증가하였다. 대조구에 비해서 75% 차광조건의 건물중이 약 2배 더 높은 값을 보였다. 건물중은 통계적으로 유의한 차이를 보였다(Fig. 3)
Table 2
The growth characteristics of Alliaria petiolata grown under three different shading treatment.
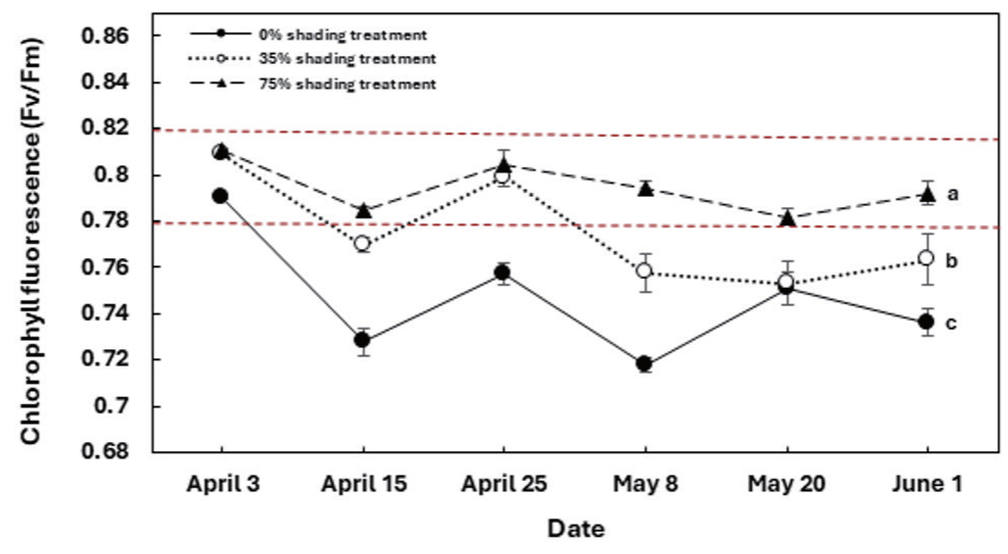
식물체에서 흡수한 광 에너지 중 광합성 초기 광화학 반응에 사용되지 않고 외부로 방출되는 광 에너지를 형광이라고 한다(Kim et al., 2018). 최대양자수율(Fv/Fm)은 최대형광에 대한 변이형광의 크기이다.(Maxwell and johnson, 2000; Kim et al., 2018). Fv/Fm은 정상적인 광합성 상태를 제공하는 것뿐만 아니라 광계Ⅱ의 광화학적 효율, 즉 스트레스 요인에 가장 민감한 매개변수를 제공한다(Kycko et al., 2018). 따라서 광합성 과정에서 엽록소 효율을 평가하기 위하여 Fv/ Fm의 값을 이용한다. 마늘냉이의 최대양자수율의 변화는 Fig. 4 와 같았다. Fv/Fm은 0%와 35%는 75%와 유의한 차이를 보였으나, 0%와 35% 사이의 통계적 유의성은 찾을 수 없었다 광계Ⅱ에서 최대양자수율을 나타내는 Fv/Fm의 변화를 보면 75% 차광조건의 경우 시험 시작과 마지막 기간까지 건강한 식물의 Fv/Fm 범위인 0.79에서 0.84 사이의 값을 가진다. 하지만 35% 차광조건의 경우 차광 처리 초반에는 0.81의 건전한 식물의 Fv/Fm값을 갖지만, 차광 처리가 길어질수록 감소하는 경향을 보였다. 차광 처리를 하지 않은 대조구 또한 4월 3일엔 0.79의 값을 보였지만, 그 후 0.79 이하의 값으로 감소하였다. 대조구와 35% 차광조건의 경우 마늘냉이는 광으로 인하여 스트레스 환경에 처해있거나, 광계Ⅱ 반응 중심이 손상을 입었다고 판단된다. 이는 낮은 엽록소 함량과 광 흡수 효율이 낮음을 의미한다(Kycko et al., 2018).
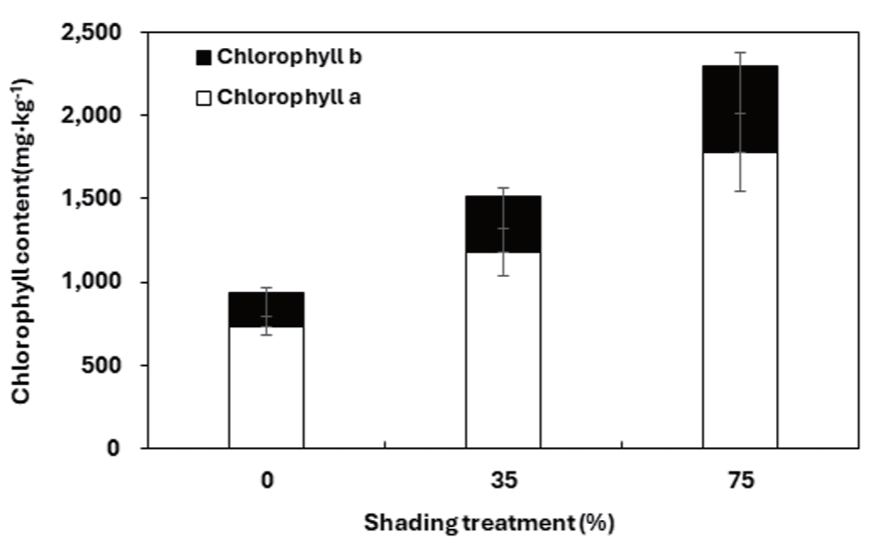
대조구 마늘냉이의 총 엽록소 함량, 엽록소 a, 엽록소 b를 측정한 결과 모두 차광 처리구에 비하여 낮은 함량을 보였다(Fig. 5). 대조구의 높은 광도 조건이 마늘냉이의 엽록소 함량을 감소시킨 것으로 판단된다. 총 엽록소 함량은 75% 차광구에서 2,292.31 mg·kg-1으로 가장 높게 나타났다. 차광율이 높아질수록 유의적 차이를 보이며 총 엽록소 함량이 증가하였다. 이와 같은 결과는 Choi and Guak (2013)에 따르면 낮은 광 조건에서 빛을 더 많이 받아들여 이용효율을 높이기 위함이라고 보고되었다. 내음성이 강한 식물이나 낮은 광도에 순화된 식물은 높은 광 조건에선 본 실험과 같은 낮은 총 엽록소 함량을 나타낸다(Je et al., 2006). 그러나 엽록소 a/b율은 대조구와 차광처리 구간별에 있어 유의적 차이를 찾을 수 없었다.
3. 차광조건별 효과적인 제거 방법 조사
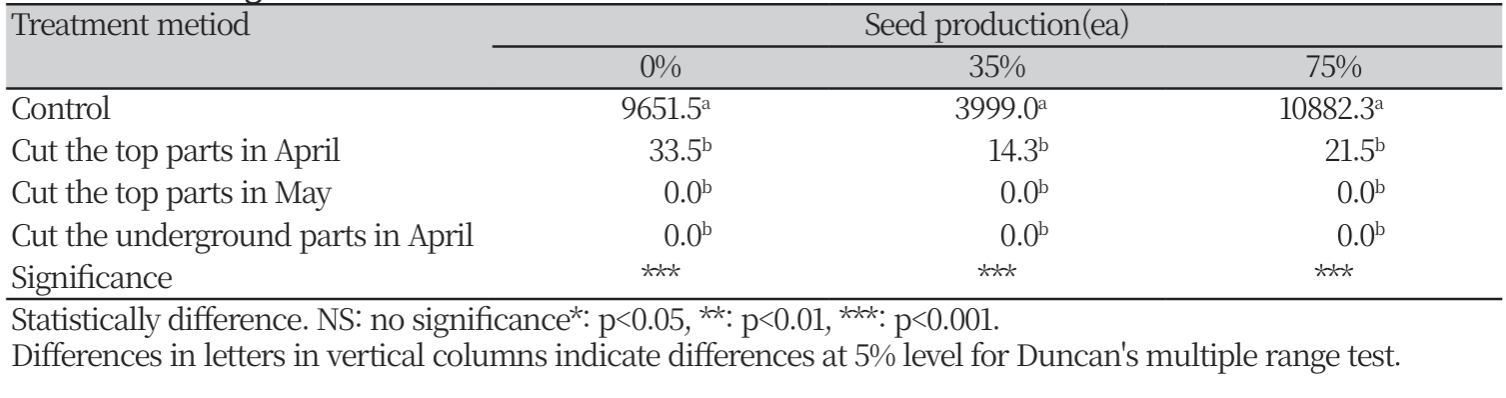
차광 처리를 하지 않은 시험구, 35% 차광 시험구, 75% 차광 시험구, 총 세 가지 차광조건에서 제거 방법에 따른 고사율을 조사한 결과, 무처리와 그 외 다른 처리방법 간의 통계적 유의성은 발견되었으나, 각 처리 방법 간의 유의적 차이는 나타나지 않았다. 차광 처리를 하지 않은 시험구와 35% 차광 처리 시험구에서는 5월에 지상부 제거와 4월에 지하부 절단 방법이 고사율 100%로 가장 처리 효과가 좋은 것으로 나타났다. 75% 차광처리 시험구에선 4월에 지하부를 절단하는 방법이 고사율 100%로 가장 처리 효과가 높게 나타났다(Fig. 6). 차광 처리를 하지 않은 시험구, 35% 차광 시험구, 75% 차광 시험구, 총 세 가지 차광조건에서 제거 방법에 따른 종자 개수를 조사한 결과, 무처리와 그 외 다른 처리 방법 사이의 유의성은 발견되었으나, 무처리를 제외한 각 처리 방법 간의 유의차는 나타나지 않았다. 모든 차광조건에서 5월 지상부 제거와 4월에 지하부를 절단하였을 경우 생산된 종자 개수는 0개로 처리 효과가 가장 높은 것으로 확인되었다(Table 3). 종자를 생산하여 번식하는 일년생 및 월년생 식물을 방제하기 위해선 해당 식물의 종자를 소멸시켜야 한다. 마늘냉이 또한 월년생 식물로 효과적인 관리를 위해선 종자 은행이 고갈될 때까지 종자생산을 방지하는 것이 중요하다(Nuzzo, 1991). 마늘냉이는 음지성 식물로 알려져있지만, 숲속과 같은 깊은 그늘부터 숲 가장자리, 산책로 및 입구와 같이 거의 완전한 햇빛을 받는 장소까지 다양한 서식지에서 발견되며(Myers and Anderson, 2003), Myers et al (2005)에 따르면 마늘냉이는 광범위한 광조건에서 적응할 수 있다고 보고되었으나, 본 연구를 통해 차광이 없는 조건에서의 생육은 국내에서 불량할 것으로 판단되나, 숲속, 숲 가장자리 등의 음지에서의 왕성한 생육과 자생식물에 대한 경쟁력을 확인하였으며, 향후 철저한 관리가 필요함을 증명하였다. 또한 방제 시기 및 방법 간 처리 효과를 규명함으로써 생태계교란 생물인 마늘냉이의 관리에 활용할 수 있을 것으로 판단된다.
요약
본 연구는 마늘냉이의 제거 및 관리 방안 수립을 위해서 필요한 기초자료를 얻기 위하여 다양한 차광조건에서 마늘냉이의 생육을 조사하였다. 그리고 광범위한 광 조건에서 효과적인 방제 방법을 찾기 위해서 각 처리를 통한 제거 효과를 규명하였다. 차광 정도에 따른 생육조사 결과, 차광률이 높을수록 초장, 엽장, 엽폭, 엽수, 엽면적, 비엽면적(SLA)이 증가하였다. 차광 처리 8주 후 대조구 대비 75% 차광조건에서 초장은 약 1.2배, 엽장과 엽폭은 2배, 엽수는 약 6배 더 높은 값을 나타냈다. 차광 처리 4주 후 엽면적과 비엽면적(SLA)은 대조구와 비교하였을 때 75% 차광조건에서 약 1.5배, 약 2배 더 높은 값을 보였다. 마늘냉이의 생체중은 75% 차광조건에서 42.06 g으로 가장 높은 값을 보였다. 건물중은 대조구와 75% 차광조건의 마늘냉이를 비교하였을 때 대조구는 15.02 g, 75% 차광조건에선 29.00 g으로 약 2배 더 높았다. Fv/ Fm의 경우, 75% 차광조건에선 생육기간 동안 건강한 식물의 범주인 0.79~0.84 사이의 값을 가졌다. 반면, 대조구는 처리 7일 후, 35% 차광조건은 처리 3주 후 0.79 이하의 값을 가졌다. 대조구 마늘냉이의 총 엽록소 함량, 엽록소 a, 엽록소 b를 측정한 결과 모두 차광 처리구에 비하여 낮은 함량을 보였다. 대조구에 비하여 75% 차광조건의 총 엽록소 함량이 약 2.4배 더 높았다. 차광조건 별 제거 방법에 따른 제거 효과를 조사한 결과, 차광 처리를 하지 않은 시험구와 35% 차광처리 시험구에서 5월에 지상부 제거와 4월에 지하부 절단 방법이 고사율 100%로 가장 처리 효과가 좋은 것으로 나타났다. 75% 차광처리 시험구에선 4월에 지하부를 절단하는 방법이 고사율 100%로 가장 처리 효과가 높게 나타났다. 생산된 종자 수량의 경우, 모든 차광조건에서 5월 지상부 제거와 4월에 지하부를 절단하였을 경우 생산된 종자 개수는 0개로 처리 효과가 가장 높은 것으로 확인되었다.

