서언
크리핑 벤트그래스(grostis palustris L.)는 다년생 잔디로서 습하고 서늘한 기후지역에 이용되는 한지형 잔디로 골프장의 그린과 페어웨이에서 가장 널리 이용되는 잔디 종 중 하나이다(Beard, 2001). 크리핑 벤트그래스는 고운 질감과 2-3 mm로 좁은 엽폭을 가지고 있으며 선명한 엽색을 나타내어 그린용 잔디로 이용되기에 적합하다(McCann and Huang, 2008). 하지만 크리핑 벤트그래스는 15-24℃의 생육적온을 가지고 있어 우리나라의 고온 다습한 기후에서는 온도 스트레스를 받고, 병원균의 번식이 증가하여 고온성 병해가 나타나기 쉽다(Beard, 2001). 또한 고온기의 경우, 잦은 강우로 인해 일정한 간격으로 예초를 수행하기 어렵고, 기계의 답압으로 잔디 생육이 불량해져 하절기에 한지형 잔디에 생장조정제를 활용하고자 하는 연구가 수행되어 왔으며(Lee et al., 2008), 고온기에서의 잔디 관리를 위해 일정한 잔디품질을 유지하고 적절한 예고관리를 통한 건강한 잔디 관리를 위해 생장조정제(plant growth regulator, PGR)를 이용하고 있다(Tae et al., 2010; Kim and Kim. 2023).
한지형 잔디와 난지형 잔디의 관리에 있어서 넓게 이용되는 트리넥사팍에틸(Trinexapac-ethyl; TE)은 잔디 생육 억제를 위해 사용하는 대표적인 생장조정제로(Fagerness et al., 2002; Lickfeldt et al., 2001; McCullough et al., 2006) 지베렐린(gibberellin, GA) 생합성 과정에서 GA20이 GA1로 전환되는 속도를 경감시켜 세포 길이 생장을 억제하여 예지 횟수를 감소시킨다고 보고되었다(Shepard and DiPaola, 2000). 잔디에서 사용되는 생장조정제는 TE 외에도 프로헥사디온칼슘(prohexadione-calcium)이나 파클로부트라졸(paclobutrazol) 등이 있고(Tae et al., 2010) 이들은 식물체내 GA 합성 속도를 늦추고 세포 신장을 억제하는 등의 생장속도 조절로 인해 여름철의 예지 횟수를 감소시키는데 주로 사용된다(Pornaro et al., 2017). 이 중에서도 일반적으로 TE는 예지 횟수를 줄여 고온기 잔디 관리 효율을 높이고, 잔디 밀도를 증대시키는 것으로 보고되었다(Hargey et al., 2016).
생육적온을 나타내는 봄철과 가을철의 한지형 잔디는 생육이 왕성하여 피해회복이 빠르지만 생육적온보다 높은 고온 다습한 시기인 여름철에는 잔디 생육과 피해회복이 느리다(Dong et al., 2022). 또한 동계 휴면기나 그린업이 이뤄지는 초봄에 답압에 의해 잔디의 손상이 발생하는 경우 잔디 생육 및 품질이 악화되어 경기력을 경감시킨다(Yu et al., 2023). 이렇듯 잔디 생육이 불량한 시기에 한지형잔디의 봄철 녹화 촉진과 잔디 회복을 위해 질소 비료 등을 살포하고 있으나 토양의 과습으로 인한 생리장해와 동절기로 인한 뿌리 생육 불량으로 잔디 회복이 어려운 실정이다(Kreuser and Soldat, 2012).
Lee et al. (2023)은 트리아졸계 농약의 과량처리에 왜화 효과를 보이고 있는 토마토(Solanum lycopersicum)에 GA를 처리했을 때 생장과 생육이 증가하고 회복된다고 보고하였다. Kang and Kim (2007)은 GA3 처리된 켄터키 블루그래스(Poa pratensis L.)와 톨훼스큐(Fesstuca arundinacea Schr.) 종자는 묘출현율이 무처리에 비해 8-9% 향상되었고 건물중, 초장 및 뿌리 생장 등 초기생육이 무처리에 비해 향상된다고 보고하였다. 이러한 GA3의 특성을 고려한다면 잔디의 회복이 필요한 곳에 활용할 수 있을 것으로 판단된다. 하지만 아직까지 GA3를 잔디에 이용한 실험과 연구는 미비한 실정이다. 따라서 본 연구는 TE 처리 후 잔디의 생육회복이 필요한 경우 식물생장촉진형 생장조정제(plant growth promoting regulator, PGPRs; GA3, GA4+7+6-BA, 6-BA)를 처리하여 생장조정제 종류별 크리핑 벤트그래스에 생육 및 품질의 변화를 조사하였다.
재료 및 방법
공시재료
본 실험은 대구대학교 과학생명융합대학 부속농장 유리온실에서 2023년 7월부터 2023년 8월까지 수행되었다. 공시 잔디는 크리핑 벤트그래스(A. palustris L.)로 품종은 ʻPenn-A1’ 품종의 종자를 A사로부터 제공받아 사용하였다. 공시 토양은 미국 골프협회(United State Golf Association; USGA)의 규격에 적합한 입경 분포를 갖는 모래는 아니었으나 극조사

각 6가지 종피 파상 방법으로 처리한 종자는 원예용 상토(바로커, 서울 바이오)를 채운 72공 다공포트에 2.5 cm 깊이로 파종하여 경기도 수원에 위치한 서울대학교 농장 내 유리 온실에서 생육하였다. 파종 후 11일 차에 개별 플라스틱 포트(75×75×80 mm)에 이식하여 파종 후 28일차까지 관찰하였다. 모든 실험은 3반복의 완전임의배치법으로 설계되었다.
발아 및 생육 조사
가시박의 발아는 파종 후 14일차까지 매일 조사하였다. 식물체의 정단부가 흙을 뚫고 표면 위로 올라온 것을 기준으로 발아 여부를 판단하였다. 가시박의 생육은 떡잎, 본엽의 엽폭 및 엽장, 엽기에 대해 7일 간격으로 조사하였다. 추가로 종피 파상이 식물의 떡잎에 주는 영향을 알아보기 위하여 파종 후 10일차에 발아한 개체 중 떡잎에 상처가 발생한 개체의 수를 조사하였다. 엽폭은 잎에서 가장 넓은 부분의 길이를, 엽장은 잎자루가 끝나는 부분에서 엽선까지의 길이를 의미한다. 엽기는 떡잎을 제외한 본엽의 마주난 잎 두 장을 한 단계로 계산하였다.
식생의 엽면적은 파종 후 18일차에 촬영한 RGB 영상에서 MATLAB R2023b 프로그램을 활용해 분석하였다(The MathWorks Inc., MA, USA). RGB 영상에서 식물체의 녹색 부분만 영역화하여 남기고, 분리된 잎의 영역의 픽셀 수를 함께 촬영한 color checker의 기준 면적과 비교하여 식물의 엽면적을 계산하였다. 또한 파종 후 28일차 생육 조사를 완료한 뒤 지상부를 수확하여 생체중을 측정하였다.
통계 분석
모든 조사 항목은 먼저 분산분석(ANOVA)을 통해 유의성을 검정하고, 유의성이 확인된 항목들은 던컨다중검정 (Duncan’s multiple range test)을 통해 통계적 차이를 비교하였다. 종자의 파종 후 일수(x)에 따른 누적 발아율(Y)은 비선형 회귀분석을 통해 수식 (1)과 같은 Gompertz 모델에 적합시켜 최대누적발아율(C), 누적발아곡선의 기울기(B), 최대누적발아율(C)의 50%에 도달하는데 소요되는 기간(M)을 구하였다.
모든 통계적 분석은 R 4.3.1 (R Foundation for Statistical Computing, Vienna, Austria)을 이용해 진행되었다.
결과 및 고찰
종피 파상 위치가 발아에 미치는 영향
파종 후 5일차에 무처리를 제외한 5가지 종피 파상 처리에서 모두 발아가 시작되었다. 최초 발아가 관찰된 파종 후 5일차에는 종피제거 처리에서 발아율이 가장 높았고, 그 뒤로 종자과반제거 > 종단제거 = 종자테두리제거 > 주공제거의 순서로 발아율이 높았다. 최대 발아율에 도달하는 시기는 종피제거가 7일로 가장 빨랐고, 무처리가 14일로 가장 느렸으며, 파상 처리 위치에 따라 발아 속도에 차이가 있음을 확인하였다. 가시박의 발아율은 종피 파상 부위의 면적이 넓을수록 빠르게 증가하였다. 파종 후 14일까지의 발아율을 Gompertz 모델에 대해 비선형회귀분석을 진행한 결과에서 B 값은 발아 속도를 나타낸다. B 값은 종피제거 > 종자과반제거 > 종자테두리제거 > 종단제거 > 주공제거 > 무처리의 순서로 높았으며, 이는 종피 파상 부위의 면적이 넓은 순서와 경향이 일치하였고, 종피 파상 면적이 동일한 종단제거 처리가 주공제거 처리보다 발아 속도가 빨랐다(Table 1). 이는 넓은 면적을 통해 물과 산소를 빠르게 흡수하여 발아가 촉진되었고, 종자의 발아 과정에서 떡잎에서의 물 흡수, 저장 양분의 소화가 먼저 진행된 후 유근의 세포 신장이 이루어지기 때문에 떡잎 쪽의 종피 파상이 발아를 더 촉진한 것으로 판단된다(Bewley, 1997).
Table 1
Parameter estimates for the Gompertz model to fit the germination rates of Sicyos angulatus with 6 different scarifications; cutting hilum area, cutting seed tip, cutting seed edge, half removal, seed coat removal, and control. The numbers in parentheses are standard errors.
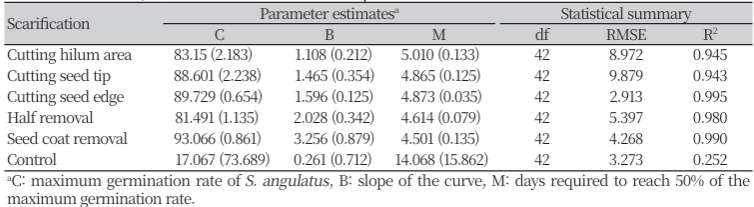
각 처리별 발아율은 발아 시작 후 초기에는 차이가 났지만, 시간이 지날수록 처리 간 차이가 줄어 파종 후 14일차의 최종 누적 발아율은 무처리를 제외하고는 유의미한 차이를 보이지 않았다. 종피 파상을 하지 않을 경우 파종 후 14일차에 약 6.7%로 낮은 발아율을 보였다. 반면 종피제거 처리는 93.3%, 종단제거와 종자테두리제거 처리는 90.0%, 주공제거와 종자과반제거 처리는 83.3%의 최종 발아율을 나타내 종피 파상으로 인한 발아 촉진 효과가 확인되었다(Fig. 2). 이러한 결과는 선행 연구의 결과와 유사하였다. Lim et al. (1994)이 가장 효과적인 파상법으로 제시한 박피 처리는 본 연구의 주공제거 처리와 동일하였다. Jones (1971)의 송곳을 이용한 종피 파상 방법과 종피제거 처리를 통한 90% 이상의 높은 발아율 확보와 이들 파상처리한 종자를 파종한 후 4-5일차에 발아가 개시되었다는 결과는 본 연구의 결과와 일치하였다.
Fig. 2
Accumulated germination rate of Sicyos angulatus until 14 days after sowing of seeds with 6 different scarifications; (A) cutting hilum area, (B) cutting seed tip, (C) cutting seed edge, (D) half removal, (E) seed coat removal, and (F) control. The red lines are fitted germination rate using the Gompertz model. The error bars represent SEM of three replicates.
종피제거, 종단제거 및 종자테두리제거 처리는 90% 이상의 높은 발아율과 함께 발아 속도 또한 빠르게 나타나, 가시박 종자의 균일한 발아 개체를 빠르게 확보하기 위한 처리 방법으로 활용성이 높다고 판단된다. 이러한 처리 방법은 선행연구의 송곳, 비커, 사포, 정미기 등을 이용한 물리적 종피 파상이나 화학적 처리(Jones, 1971; Lim et al., 1994; Mann et al., 1981)에 비해 비교적 간편하며, 높은 발아율 확보가 가능하였다. 따라서 종피 파상 위치를 달리한 본 연구의 파상처리 방법은 가시박의 균일한 발아가 필요한 연구에서 실용적으로 활용 가능할 것으로 사료된다.
종피 파상 위치가 초기 생육에 미치는 영향
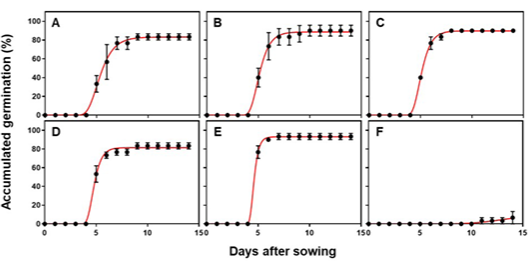
파종 후 10일차에 가시박의 떡잎을 관찰한 결과 주공제거 처리에서는 떡잎에 상처를 입은 개체가 발생하지 않았다(Fig. 3). 그러나 종자테두리제거 처리에서는 약 20%의 개체에서 떡잎에 상처가 발생하였고, 종단제거와 종피제거 처리에서 약 50%의 떡잎에 상처가 발생하였다. 종자과반제거 처리는 모든 개체의 떡잎에 상처가 발생하였다(Fig. 4A).

파종 후 18일차에 가시박의 엽면적을 측정한 결과 떡잎의 절반을 제거한 종자과반제거 처리의 엽면적이 매우 낮은 결과를 보였다. 이는 발아 후 초기 생육에 필요한 양분을 떡잎에 저장하는 박과 식물의 특성 상 떡잎의 많은 손상이 양분 부족으로 이어진 것으로 보인다. 하지만 종자과반제거 처리를 제외한 나머지 처리구에서는, 떡잎의 손상 여부보다는 발아 속도의 차이가 초기 생육 단계의 진전 속도에 더 주된 영향을 미친 것으로 확인되었다(Fig. 4B).
Fig. 4
Damaged cotyledon (%) (A) out of emerged seedlings at 10 days after sowing and leaf coverage (cm2, B) of Sicyos angulatus at 18 days after sowing of seeds with 6 different scarifications; cutting hilum area, cutting seed tip, cutting seed edge, half removal, seed coat removal, and control. The error bars represent SEM of three replicates.
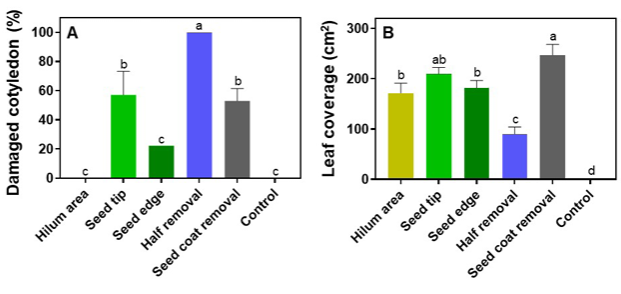
가시박의 엽기 전개속도에 있어 생육 초반에는 각 처리 별로 차이가 있었다. 엽기는 엽면적 측정 결과와 비슷한 경향으로 종피제거 처리에서 엽기가 가장 빠르게 전개되었고, 그 뒤로 종단제거, 종자테두리제거, 주공제거 처리가 빨랐고, 종자과반제거의 엽기 전개가 가장 느렸다. 그러나 이러한 경향은 파종 후 28일차까지 생육이 진전됨에 따라 차이가 줄어들었으며, 특히 종단제거, 종자테두리제거 처리의 엽기 전개는 파종 후 28일차 조사에서 종피제거 처리와 유의한 차이를 보이지 않았다(Fig. 5A).
파종 후 28일차에 조사한 가시박의 생체중 또한 다른 생육 지표들과 동일하게 종자과반제거 처리와 무처리에서 낮았으며 이를 제외한 4가지 처리 간의 유의한 차이가 없었다(Fig. 5B).
본엽의 엽폭과 엽장을 파종 후 28일차에 조사한 결과 종자과반제거 처리를 제외한 모든 처리는 모든 1, 4, 8엽에서 유의한 차이가 없었으며, 종자과반제거 처리 1엽의 엽폭과 엽장이 모두 다른 처리와는 통계적으로 유의한 수준으로 작았다. 보다 생육이 진행된 뒤에 전개된 4엽과 8엽에서는 종자과반제거 처리와 다른 처리 간의 차이가 줄어들었지만 유사한 경향을 보였다. 실험 결과를 통해 본엽의 엽폭 및 엽장의 경우 떡잎의 손실로 인한 차이를 제외하고는 처리 간 큰 차이가 없음을 확인하였다(Fig. 5C, 5D). 낙엽성 관목 Prosopis juliflora의 종피 파상 연구에서, 사포를 이용한 물리적 종피 파상 방법은 발아 촉진 효과가 우수했으나 떡잎 손상으로 인해 기형 묘목의 발생이 증가하고, 생존율은 감소한다고 보고된 바 있다(Miranda et al., 2011). 본 연구의 50% 종자를 제거한 종자과반처리에서 확인된 생육 저하도 떡잎의 과도한 손실이 초기 생육에 영향을 미쳤기 때문으로 판단된다.
Fig. 5
Leaf number (A) of Sicyos angulatus measured at 14, 23, and 28 days after sowing, fresh weight (B) at 28 days after sowing, widths (C) and lengths (D) of the 1st leaf, 4th leaf, 8th leaves measured at 28 days after sowing of seeds with 6 different scarifications; cutting hilum area, cutting seed tip, cutting seed edge, half removal, seed coat removal, and control. The error bars represent SEM of three replicates.
종합하면, 초기생육 조사 결과, 생육 초반에는 종피제거 처리가 빠른 발아 속도를 통해 상대적으로 빠르고 균일한 초기 생육을 보였으나, 생육이 진행됨에 따라 점점 처리 간의 차이가 줄어드는 것을 확인하였다. 반면 종자과반제거 처리는 떡잎의 손실로 인해 발생한 초기 생육을 회복하지 못하여 지속적으로 부진한 생육을 보였다(Hanley and Fegan, 2007). 가시박을 실험 목적으로 파종할 경우 발아 및 생육의 균일성이 중요한데, 본 연구 결과 종피 파상 면적이 넓을수록 발아 과정 초기에 수분을 빠르게 흡수하여 발아 속도가 빠르고, 이후 초기 생육 또한 균일하게 진행되는 것을 확인하였다. 이는 지상 발아의 특성을 가진 가시박에서 발아 이후 전개되는 떡잎이 바로 광합성 및 동화작용을 시작하기 때문에 떡잎의 손상보다는 발아의 속도가 큰 영향을 끼친 것으로 보인다(Hanley et al., 2004). 본 연구에서 평가한 여러 파상처리 방법 가운데 종피제거 처리가 가장 효과적이나 종피제거를 위한 세심한 주의와 시간이 많이 요구된다. 따라서 종피제거 처리 보다 방법적으로 간편한 종자테두리제거 처리 혹은 종단제거 처리는 균일한 높은 발아와 초기생육을 확보하는 데 현실적인 대안이 될 수 있다. 본 연구 결과는 가시박 종자의 균일한 발아와 초기 생육 확보를 위한 종피 파상 처리 기준을 제시하였으며, 향후 가시박의 생리생태적 연구나 방제관리 방법 마련을 위해 가시박을 재배하고자 할 경우에 활용될 수 있을 것으로 기대된다. 또한 가시박 외에도 박과 식물 등 물리적 휴면을 갖는 대립 경실 종자의 휴면을 타파하는 데 본 연구에서 제시한 방법을 적용할 수 있을 것으로 판단된다.
요약
가시박(Sicyos angulatus)은 종피의 불투수성으로 인한 물리적 휴면을 가지고 있어, 연구 목적의 균일한 발아를 유도하기 위해서는 인위적인 종피 파상 처리가 필요하다. 본 연구에서는 물리적 종피 파상 위치가 가시박의 발아 및 초기 생육에 미치는 영향을 구명하기 위해 수행되었다. 실험 결과, 종피 파상 면적이 넓을수록 발아 속도가 빨라지는 경향을 보였다. 파종 후 발아가 가장 빠르게 진행된 종피제거 처리가 생육에 있어서도 가장 빠른 양상을 보였으며 종단제거와 종자테두리제거 처리가 양호한 초기 생육을 보였다. 종자과반제거 처리는 넓은 종피 파상 면적으로 인해 발아 속도는 빨랐으나, 떡잎의 심각한 물리적 손상으로 인해 초기 생육이 지연되는 양상을 보였다. 파종 후 28일차의 최종조사에서 종자과반제거 처리를 제외한 모든 종피 파상 처리 간의 생육(엽기, 엽폭, 엽장, 생체중)에 유의미한 차이가 없었으며, 종자과반제거 처리의 경우 떡잎 손상으로 인한 초기 생육 부진을 본엽이 전개되어도 회복하지 못하였다. 결론적으로 가시박의 제초제 스크리닝 등 균일한 식물체 확보가 필요한 연구에서는 발아가 빠르고 균일한 종피제거 처리가 가장 효과적이나, 작업 효율성을 고려한다면 종자테두리제거 또는 종단제거 처리가 유용한 가시박 종피 파상법으로 활용될 수 있을 것으로 판단된다.

