서 언
한국잔디가 1900년대 초반 미국으로 도입된 이후 잔디로서의 가치는 1970년대 중반부터 미국을 중심으로 크게 인정되기 시작하였다(Patton et al., 2017). 에너지의 부족, 노동력의 경제적 부담 증가, 관리효율의 경제성은 최근 환경보전의 특별한 관심과 함께 물 등 자원 투입이 적고, 환경 적응성이 우수한 잔디 품종의 개발 필요성이 대두되었다.
최근 기후변화의 영향으로 인해 한반도 기후 또한 점점 아열대화 되어가고 있어 이러한 기후변화 적응성이 높은 난지형 잔디인 한국잔디 품종 개발이 요구되고 있다(Feng et al., 2019).
한국잔디는 우수한 내환경성과 환경보호적 기능성이 우수하고(Emmons, 1995), 거의 모든 기후와 토양에서 생장할 수 있기에 공원, 골프장, 제방, 운동 경기장에서 넓게 사용되고 있다(Al-Khayri et al., 1989; Patton et al., 2017). 한국잔디의 신품종 육종을 위한 주된 목표는 녹색기간이 연장되는 품종, 내답압성이 우수하여 스포츠용으로 활용 가능한 품종, 내염성 및 내건성 품종, 관리요구도가 낮은 내척박성 품종 등이 제시되고 있다(Joo et al., 1997).
건조와 염 등은 환경스트레스의 중요한 원인 중의 하나로 세포내에 유해한 활성산소종(reactive oxygen species, ROS)들이 생성되어 엽록체나 미토콘드리아의 전자전달을 저해하고, 엽록소 함량과 성분의 변화와 광합성 기구의 손상으로 인해 식물의 광합성 능력이 감소된다(Lee et al., 2008; Miller et al., 2010). 또한 지질 과산화로 인한 세포막 파괴, 세포의 구성성분 및 단백질 변화 등 산화적 손상에 대응하는 항산화 방어 체계가 파괴되어 산화 스트레스로 인해 생육이 억제된다(Fu and Huang, 2001; Iturbe-Ormaetxe et al., 1998).
이러한 산화적 손상으로부터 잔디는 항산화물의 축적 또는 항산화 효소에 의한 산화물질들의 소거 기능을 통해 스트레스로 인한 세포 또는 세포막 등의 피해로부터 여러가지 생리적 대사 활동을 유지하게 되고, 이러한 특성은 한지형 잔디 종류 간에 차이가 발생한다는 것이 알려졌다(Jiang and Huang, 2001). 수분 부족과 높은 염도 환경에서 자란 한지형 또는 난지형 잔디는 산화력이 높은 활성산소종의 증가가 높았고, 이러한 활성산소종의 증가로 세포막을 구성하는 지질의 산화가 두드러지게 나타나고 있다고 보고하고 있다(Du et al., 2009). 토양 수분의 절대적 부족에 의한 건조 스트레스와 염분 함량이 높은 토양 또는 수질에 의한 염분 스트레스와 같은 열악한 환경에서 잔디의 품질 저하는 공통적으로 뿌리 발달의 감소, 낮은 식물체 내 수분 포텐셜 유도, 광합성 및 광합성 산물의 축적 저하와 관련이 있다(Carrow, 1996; Lee et al., 2004). 따라서 내건성 또는 내염성이 있는 품종은 우수한 항산화 능력을 통해 생리적 대사작용을 유지하고, 특히 산화물에 의한 세포 인지질 막의 손상으로부터 그 기능이 상대적으로 높게 나타나고 있음이 보고되고 있다(Jiang and Huang, 2001).
내건성 또는 내염성 기작은 여러 가지 요인이 작용하기 때문에 후보 유전자원의 선발이 쉽지 않다. 하지만 수분 스트레스 또는 높은 염분 토양 환경에서 높은 지상부 생육을 보이는 개체를 선발하게 되면 이러한 스트레스 환경에서 내성을 가진 1차 후보 자원이 될 수 있으며, 산화물에 의한 세포의 보호 기능과의 상관성을 살펴본다면 향 후 내건성 또는 내염성 육종에 많은 기여를 할 수 있을 것이다. 따라서 본 연구는 교배를 통해 만들어진 한국잔디 교배후대에 대한 특성 조사와 산화물로부터 세포막의 보호 기작을 기초로 건조와 염 저항성 후보를 탐색하고, 향 후 이러한 단일 또는 복합 환경스트레스 조건에서 내환경성 품종을 육성하고자 유전자원 스크리닝 연구를 수행하였다.
재료 및 방법
한국잔디의 인공교배
2010년 5월부터 2011년 9월까지 남해안, 동해안 및 서해안 도서, 내륙과 산간지역에서 242개체의 한국잔디 유전자원을 수집하였다. 수집된 유전자원은 경남 진주시 소재의 국립산림과학원 산림바이오소재연구소 잔디유전자원 보존원에 이식을 하였다. 1.3 m×1.3 m 크기의 시험구에 마사토를 깔아 수집해 온 잔디를 이식한 후 증식을 시켰다. 초기 증식 기간동안 수집지에서 가져온 상태를 그대로 유지하기 위해 잔디 깎기와 비료를 시비하지는 않았다. 2012년 2월부터 2013년 4월까지 잔디유전자원 보존원에 식재 된 242개체를 대상으로 형태적 특성, 피복 상태, 봄철 그린업, 화서 발생량 등 특성조사를 수행하였다. 엽폭이 중·세엽형, 빠른 생장, 높은 밀도, 화서 발생량이 많은 특성을 보이는 교배 후보 36개체를 선발하였다. 선발된 교배 양친은 2013년도 5월에 21.5 cm×21.5 cm×8cm 크기의 화분에 마사토를 채운 후 지름 10 cm 홀컷트로 보존포에서 증식하고 있는 잔디를 채취하여 옮겨 심었다. 식재 후 온도 최저 10℃, 최고 35℃, 습도 최저 40%, 최고 70% 온실조건에서 생육을 시켰다. 2014년 4월부터 선발된 유전자원과 상업종(제니스)을 활용하여 101개의 교배조합을 작성하고 인공교배를 수행하였다. 5월부터 암술과 수술이 올라오기 시작하면 출현되는 꽃대에 유산지 봉투를 씌우는 방식으로 인공교배를 수행하였다. 한국잔디는 자예선숙의 특성(Choi et al., 2008; Forbes, 1952)이 있기 때문에 암술만 개화된 상태에서 제웅없이 꽃가루를 받아 인공교배를 수행하였다. 인공교배 1개월 후 꽃대를 잘라 유산지 채로 채취하였으며, 종자 후숙을 위해 종자 채취 후 약 30일 동안 실온 상태에 두었고, 후숙 된 종자는 4℃ 냉장고에 보관하여 후대 개체 증식을 위한 파종에 사용하였다. 2014년 8월에 종자를 25% KOH에 30분간 침지 후 흐르는 물에 12시간 수세하여 페트리디쉬에 파종하였다. 파종한 종자는 식물생장상(DS52G4P, Dasol Scientific Co., LTD., Hwaseong, Korea)에서 발아를 시켰으며, 식물생장상 환경은 20℃, 16시간 암조건, 35℃ 8 시간 광 조건, 습도 60-80% 조건이었다. 발아된 개체는 상토를 채운 플러그 트레이에 이식을 하여 교배후대(F1) 522개체를 획득하였다. 획득한 교배후대 개체는 온실에서 생육을 시킨 후 2015년도에 1차로 빠른 피복력과 생육이 우수한 60개 계통을 선발하였고, 선발된 계통들은 마사토를 채운 삽목 상자에 이식을 하여 증식을 시켰다. 2016년도에는 선발된 60개 계통의 특성평가를 수행하여 중·세엽형의 엽폭을 보이면서 지상부 줄기 밀도가 높은 32개 개체를 선발하였다(Table 1). 형태적 특성은 초장, 엽장, 엽폭, 최하위 잎 높이를 조사하였으며, 생육 속도가 비슷한 줄기 10개를 무작위로 선택하여 측정한 후 평균 및 표준오차를 계산하였다. 초장은 지면으로부터 가장 높게 자란 잎까지의 높이를 측정하였고, 엽장은 기엽(1번째)으로부터 3번째 잎의 길이를 조사하였고, 엽폭은 3번째 잎의 최대 엽폭으로 자를 이용하여 mm이하 한자리까지 측정하였다. 지면에서부터 첫째 잎까지의 높이는 지면으로부터 줄기 첫째 붙은 잎의 기부까지의 높이를 측정하였다(Choi et al., 2012). 지상부 줄기 밀도는 5 cm×5 cm 크기의 사각 틀을 이용하여 4반복으로 사각틀 내의 지상부 줄기의 개수를 측정하였다.
|
Table 1. Morphological and growth measurements of selected 32 breeding lines out of initial 522 zoysiagrass hybrids over two years. 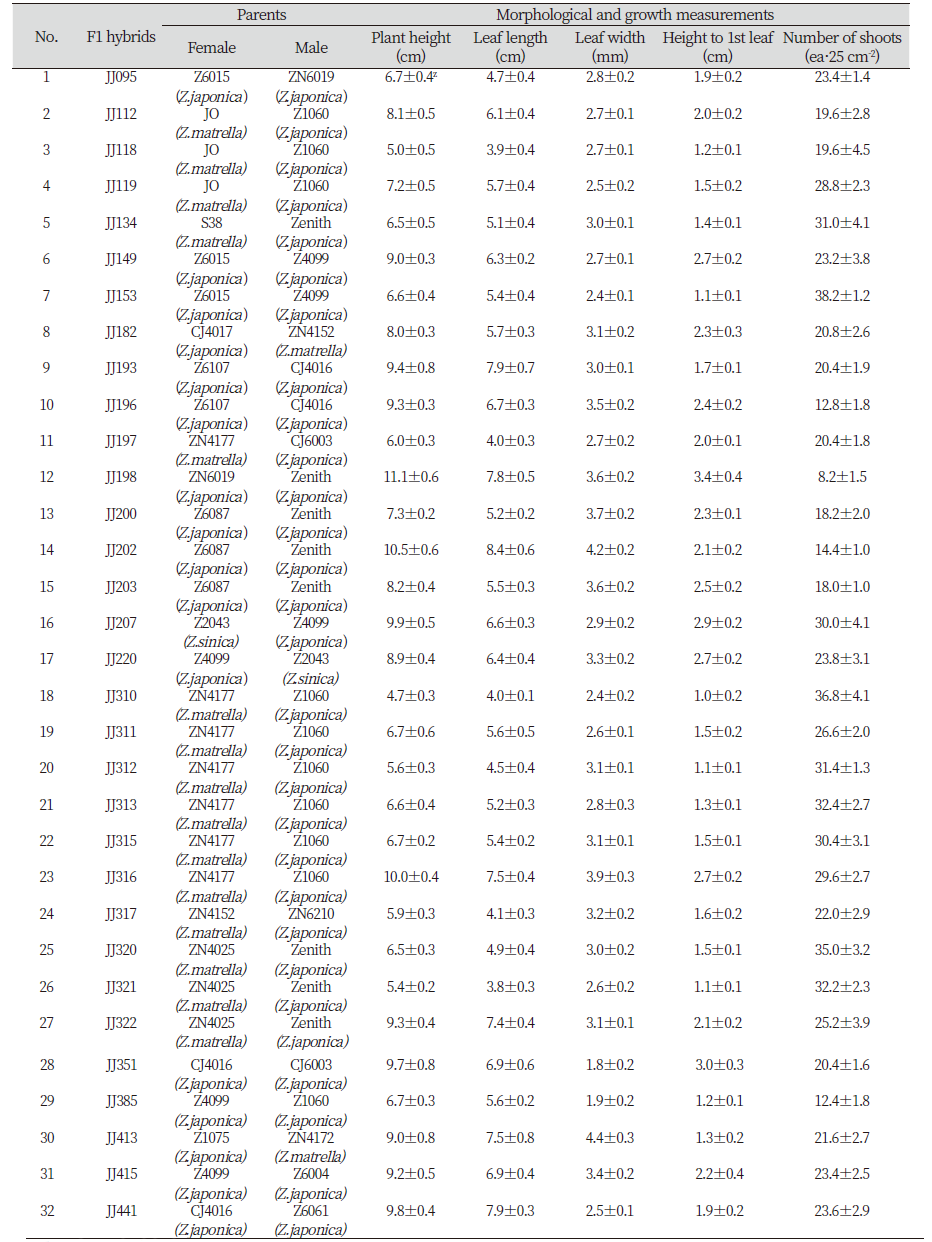
|
|
z Mean ± standard deviation. |
건조 및 염 스트레스 처리 후 세포막 지질 과산화성 간접 검정
선발된 교배후대 32개 후대 개체들의 건조와 염스트레스에 대한 차이를 알아보기 위해 스트레스 지표인 지질의 과산화(malondialdehyde, MDA) 함량을 분석하여 수분과 염 스트레스에 대한 내성 평가를 수행하였다.
2017년 6월 선발된 교배후대는 지하경과 포복경을 채취하여 지름 12 cm 화분에 마사토를 채운 후 각 화분 당 25 cm씩 지하경과 포복경을 식재하였다. 식재 후 유리온실에서 생육을 시켰으며, 관수는 1일 1회 두상관수를 하였으며, 관비는 Hoagland 양액을 2주에 1회씩 공급하였다.
2017년 9월 생육 된 교배후대들은 식물생장실(plant growth room)로 옮겨 건조와 염스트레스에 대한 저항성 검정을 수행하였다. 생장실의 환경조건은 온도 26-28℃, 광조건 명기 12시간, 습도 50-60%를 유지하였다. 건조 처리구는 12일간 관수를 하지 않았고, 염 처리구는 0.6% NaCl을 2일 간격으로 20일 동안 두상관수를 하였다. 실험구는 8반복 완전임의 배치법으로 설계하였으며, 건조 처리 12일 이후 토양수분측정기(HMM-100Pro, Hanyoung system, Seoul, Korea)로 토양수분을 측정하여 수분 장력 0 pF를 유지하지 않은 8개(JJ153, JJ182, JJ193, JJ196, JJ197, JJ203, JJ310, JJ311) 계통들을 제외한 나머지 24개 계통과 염 처리 20일 후 32개 계통들의 환경스트레스에 대한 세포막의 안정성과 과산화물질을 알아보기 위해 MDA 함량을 측정하였다.
불포화지방산의 분해산물인 MDA 함량은 Heath and Pacher (1968)의 방법에 준하여 채취한 잎 생체시료 0.2 g을 5 mL의 5% trichloroacetic acid (TCA) 용매에 균질화한 후 20분간 4℃에 12,000×g 원심분리(5810R, Eppendorf, Hamburg, Grmany)하였다. 상층액 2 mL에 같은 양의 2 mL 0.6% thiobarbituric acid (TBA) 용매를 첨가한 뒤 15분 동안 80℃ 항온수조에서 반응을 시켰다. 추출 용매를 다시 4℃에 10분간 12,000×g로 원심분리한 후 분광광도계(UV-1800, Shimadzu, Kyoto, Japan)로 450, 532, 600 nm의 파장에서 흡광도를 측정하여 다음 식에 대입하여 계산하였다.
MDA (nmol·L-1)=6.45×(A532-A600)-0.56×A450 (1)
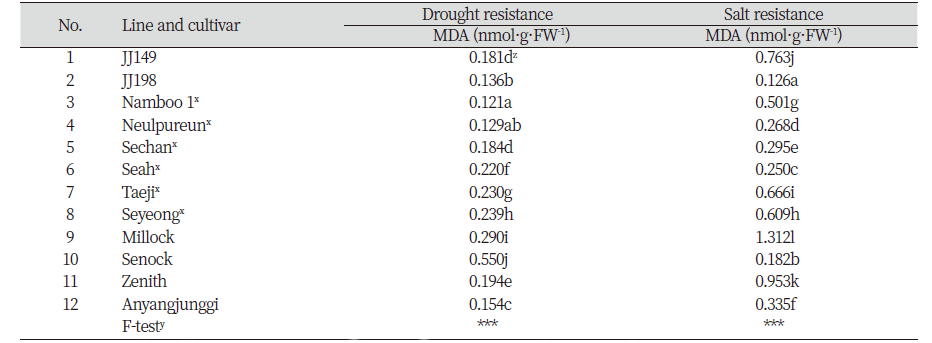
2018년 직전 2017년 조사에서 건조와 염 스트레스에 대한 내성이 우수한 2개 선발 계통과 산림바이오소재 연구소에서 품종보호 출원한 남부1, 늘푸른, 세찬, 세아, 태지, 세영 등 6 개 품종, 그리고 기존 상업 품종 안양중지, 세녹, 밀록, 제니스 등 4 품종을 상기와 같은 방법으로 건조와 염 저항성 비교를 위해 MDA 함량을 통한 간접 검정을 다시 한번 실시하였다.
통계분석
통계분석은 SAS 프로그램(v. 9.1, Cary, NC, USA)을 사용하여 처리구별 평균간 유의성 검정을 위하여 Duncan의 다중검정 방법을 5% 수준에서 실시하였다. 또한 조사한 항목 간의 상관성과 상관계수(r)의 유의성 검정을 위하여 Pearson correlation 분석을 실시하였다.
결과 및 고찰
본 연구에서 특성 조사에 이용된 교배후대 개체들은 국내에서 수집된 242개 수집종 중에서 한국잔디의 육종 목표 형질 중에서 세엽, 재생 속도, 밀도 또는 종자 생산량에 따라 교배양친으로 36개 후보 수집종이 결정되었다. 또한 이들과 상업 품종 ‘제니스’와 교잡으로 얻어진 522 개체로부터 2년 간(2015, 2016년) 지상부 형태 및 생육 특성 평가 후에 선발된 최종 32개 교잡종들이다(Table 1). 교배 종자친 또는 화분친으로 이용된 한국잔디는 들잔디, 금잔디 그리고 갯잔디 등 3종(species)이 포함되었다. 조사에 포함된 총 32개 교잡종 중에서 교배 양친으로 금잔디는 17개 교배조합에 이용되었고, 2개 조합에서는 갯잔디가 이용되었고 나머지 13개 교배조합은 양친이 모두 들잔디였다.
생육속도와 관련된 초장 및 지상부 줄기 밀도 조사에서는 각각 4.7-11.1 cm와 8.2-38.2 개·25 cm-2의 범위의 값을 보였다. 일반적으로 금잔디를 종자친으로 이용된 교배 후대 개체에서 들잔디 또는 갯잔디와의 교잡 후대들보다 초장이 짧은 것을 알 수 있었다. 그리고 반대로 밀도는 금잔디 후대에서 상대적으로 높은 경향을 보였다(Table 1). 잎의 형태적 특성과 관련된 엽장과 엽폭 측정값은 각각 3.8-8.4 cm와 1.8-4.4 mm의 범위에서 조사되었다. 국내 주요 잔디 재배 생산지인 장성지역에서 수집한 101 수집종의 경우와 비교해 볼 때 재배지 수집종들은 엽장과 엽폭 조사에서 각각 평균 14.1 cm와 3.4 mm를 보여 32개 교배후보들보다 잎이 상대적으로 길었다는 것을 알 수 있었다(Choi et al., 2012). 이러한 차이는 장성지역 잔디 생산지역에서는 일부 금잔디 계통의 품종도 재배되고 있지만 대부분 절간 길이가 길고, 생육속도가 빠른 중지류(Zoysia spp.)에서 유래한 자원들이라는 특징 때문으로 보여 진다.
예초 높이를 결정하는데 중요한 특성으로 여겨지는 지면에서부터 첫 번째 잎까지의 높이에서는 1.0-3.4 cm 범위에서 조사되었다(Table 1). 금잔디와의 교배 후대 개체들에서 평균적으로 낮은 경향을 보였고, 갯잔디와의 교배후대 개체들(JJ207, JJ220)에서 높았다. 그리고 들잔디끼리의 교배후대 중 하나인 JJ198에서 최대값을 보였다. 잔디의 경우 줄기 위 마디에서 새로운 잎이 발생하기 때문에 줄기 절간길이는 다른 식물에서와 같이 지베렐린의 합성과 관련이 있을 것으로 추측된다. 깊은 물과 낮은 물에서 자라는 두 가지 다른 벼(Oryza sativa) 품종의 경우 절간 길이가 긴 깊은 물 적응의 품종은 절간 길이는 영양 및 생식 생장단계와 무관하게 생육 전 주기에 길이 지속적인 생장을 보였고, 외부에서 살포된 지베렐린에 대하여 훨씬 민감하고 높은 반응을 보였다고 보고되고 있다(Nagai et al., 2020). 따라서 지베렐린 처리 후에도 절간 길이 생장이 낮거나 반응이 둔감한 한국잔디 교잡종 또는 자연 교배 후대 개체들을 선발하는 것은 예초 저항성 품종의 개발에 활용될 수 있는 중요한 지표가 될 수 있을 것으로 판단된다.
잔디의 지상부 줄기 밀도는 25 cm2 면적에서 조사한 신초의 숫자로 실험에 이용된 총 32개체로부터 8.2-38.2개 범위로 교잡 계통 간에 매우 큰 차이를 보이고 있음을 알 수 있었다(Table 1). 피어슨 상관계수 분석 결과 잔디의 밀도는 초장, 엽장, 엽폭, 그리고 지면에서부터 첫째 잎까지의 높이와 각각 r=-0.48, -0.41, -0.23, -0.50으로 모두 부(-)의 상관성을 보였고, 통계적으로도 엽폭을 제외하고 모두 유의적으로 상관성을 보이고 있음을 알 수 있었다(Table 2). 즉 생육속도가 빠르고 절간 길이가 길수록 지상부 줄기 밀도는 감소한다는 것을 보여주고 있다. 이러한 사실은 특히 지면에서부터 첫째 잎까지의 높이가 높아 예초 저항성이 낮을 것으로 판단되는 들잔디 간의 교배종 JJ198과지면에서부터 첫번째 잎까지의 높이가 1.1±0.1 cm로 낮아 예초 저항성이 높을 것으로 판단되는 들잔디 간의 교배종 JJ153의 경우 에서와 같이 지상부 줄기 밀도가 각각 최소값(8.2±1.5 개·25 cm-2)과 최대값(38.2±1.2 개·25 cm-2)을 보여 서로 대조적인 결과를 나타내고 있음을 통해서도 잘 알 수가 있었다(Table 1). 잔디처럼 화본과 작물인 벼의 경우도 분얼수와 식물체 크기는 부(-)의 상관성을 보이는 것으로 알려졌는데, 잔디의 경우 밀도는 예초 빈도 등 관리방법에 의한 영향도 있지만(Hull, 2000), 벼의 경우에는 높은 유전력을 보이는 양적 형질임이 알려졌고 이들 여러 유전자는 생육단계에 따라 다르게 영향을 주고 있다고 보고되었다(Cui et al., 2004).
|
Table 2. Pearson correlation coefficients (r) among growth parameters of 32 selected progenies. 
|
|
NS: Not significant. *, **, ***: significant at 0.05, 0.01, and 0.001 probability level, respectively. |
건조와 염 등은 환경스트레스의 중요한 원인 중의 하나로 세포내에 유해한 활성산소종을 생성하여 산화스트레스를 야기한다(Lee et al., 2008; Miller et al., 2010). 한편 내건성 또는 내염성 잔디 품종들은 산화물에 의한 세포막의 손상으로부터 보호하는 기능이 우수한 것으로 알려졌다(Jiang and Huang, 2001). 한지형 잔디 켄터키 블루그래스와 톨 훼스큐의 경우 산화적 스트레스로 항산화 효소의 활력이 감소하면서 세포막 인지질 과산화가 발생하면서 동시에 MDA 함량이 증가하는 것으로 알려졌다(Jiang and Huang, 2001).
따라서 건조와 염 스트레스에 대한 저항성 개체를 스크리닝하기 위하여 MDA 함량을 분석하였다(Table 3 and Table 4). 건조 스트레스 처리 후 교배후대 24계통 간의 MDA 함량을 분석한 결과 JJ149, JJ198, 그리고 JJ134가 각각 0.04, 0.05, 그리고 0.06 nmol·g·FW-1로 통계적으로 유의적인 차이를 보이며 가장 낮은 결과를 보였다.
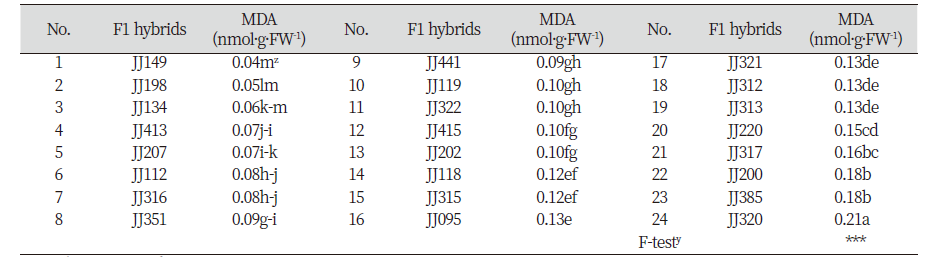
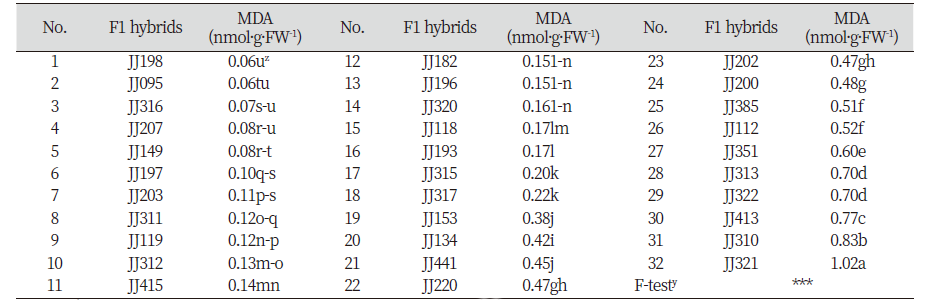
염 스트레스 처리 후 교배후대 32계통 간의 MDA 함량을 분석한 결과 통계적으로 JJ198, JJ095, JJ316, 그리고 JJ207이 각각0.06, 0.06, 0.07, 0.08 nmol·g·FW-1로MDA 함량이 유의성 있게 낮은 함량을 보여 염에 저항성이 높은 계통으로 선발되었다. 그러나 예상과 달리 건조 스트레스와 염 스트레스 처리 후 MDA 함량 간의 상관계수(r)는 0.11로 서로 유의적으로 관련이 없는 것으로 분석되었다(자료 미제공). 또한 본 연구소에서 출원한 계통 2 개체와 기존 6개의 상업품종과의 비교 실험에서 1차 조사에서 내건성과 내염성이 있을 것으로 선발된 JJ149와 JJ198 계통의 MDA 함량은 각각 건조 처리시 4.5배와 2.7배, 그리고 염 처리 시 9.5배와 2.1배 차이를 보여 MDA 함량에 큰 차이가 있음을 알게 되었다(Table 5). 이러한 사실을 통해 내건성 및 내염성 검정 시 MDA 함량만으로는 저항성 계통의 선발에 한계가 있다고 여겨지므로 건조 또는 염 스트레스 시 저항성 기작과 관련된 다른 생리적 또는 분자적 지표 중에서 표현형과 상관성이 높은 특성을 함께 분석하는 것이 바람직 할 것으로 판단된다(Fig. 1).

Authors Information
Eunji Bae, https://orcid.org/0000-0003-4597-8873
Chungyeol Kim, Forest Biomaterials Research Center, National Institute of Forest Science, Postmaster Researcher
Junhyuck Yoon, Forest Biomaterials Research Center, National Institute of Forest Science, Researcher
Eonju Jin, Forest Biomaterials Research Center, National Institute of Forest Science, Postdoctoral researcher
Youngsun Kim, Division of Life & Environmental Science and Institute of Basic Science, Daegu University, Professor
Geungjoo Lee, Department of Horticulture and Department of Smart Agriculture Systems, Chungnam National University, Professor

