서 언
유전자변형 작물(Genetically modified, GM)은 1996년 최초로 상업화된 이후 지속적으로 재배면적이 확대되고 있다. 2019년에는 1억 9,040만 ha에서 GM작물이 재배되었으며, 전 세계적으로 재배되는 면화의 79%, 콩의 74%, 옥수수의 31%, 유채의 27%가 GM작물이었다(ISAAA, 2019). 이에 따라 GM작물의 종자 시장 규모도 꾸준히 성장하여 2021년에는 약 197억 달러로 전체 종자 시장의 35.8% 가량을 차지하는 것으로 추정되었다(OECD, 2018; The business research company, 2022). 전 세계적으로 GM작물의 재배 및 유통 규모가 증가함에 따라 GM작물의 환경 방출 사고 역시 증가하고 있다 (Price and Cotter, 2014).
우리나라는 아직까지 GM작물의 상업적 재배가 승인된 바 없으나 식품·사료·산업용으로 수입하는 GM작물의 규모가 꾸준히 증가하여 최근에는 매년 1천만 톤 이상을 수입하고 있으므로(KBCH, 2021), GM작물의 환경방출에 의한 도입유전자의 생태계 침입 우려에서 자유롭지 못하다. 수입된 GM작물이 운송 중 낙곡되어 비의도적으로 환경방출되는 사례가 수차례 보고되었으며(Kim et al., 2006; Kim et al., 2016; Lee et al., 2009a), 2017년 전국적 규모로 발생한 미승인 GM유채 및 GM면화의 환경방출사고를 계기로 GM작물의 환경위해성에 대한 우려가 급속도로 높아지고 있다. 특히 우리나라의 농업생태계는 자연생태계와 경계가 명확하지 않고 서로 인접한 경우가 많아 GM작물의 환경방출에 따른 생태계 교란과 이로 인한 생물종 다양성 감소 피해가 클 것으로 예상된다(Yook et al., 2021).
GM작물의 환경방출 시 생태계에 대한 가장 큰 위해 요소는 도입 유전자(transgene)의 이동으로 화분 또는 종자의 형태로 발생할 수 있다. 종자 상태로 방출된 GM작물은 재배종 특성상 낮은 휴면성과 높은 양분 요구도 등을 갖고 있어 인위적인 관리가 배제된 자연 생태계 내에서의 위해성은 일반적으로 낮다. 그러나 화분비산에 의해 GM작물의 도입유전자가 근연 야생종 또는 잡초종으로 확산될 경우, 교잡후대종이 높은 적응성(fitness)을 획득하여 생태계 내 지속 가능성이 높아진다(Liu et al., 2013). 또한 GM작물의 도입형질은 선택압력이 존재할 경우 생태계 내 지속성을 크게 높이게 된다(Mason et al., 2013; Londo et al., 2010). 따라서 본 리뷰논문에서는 세계 4대 GM작물인 콩, 옥수수, 유채 및 면화와 국내 주곡 작물인 벼를 대상으로 현재까지 재배 승인된 형질전환 이벤트의 도입형질 정보, 국내 생태계에 분포하는 자연교잡 가능 근연종과 국내 환경조건에서 수행된 유전자이동성 연구 결과 등을 종합하여 이들 GM작물들과 근연종 간의 교잡가능성을 논하고자 한다.
주요 GM작물의 도입형질
국제생명공학응용정보서비스(ISAAA)의 데이터베이스에 따르면, 주요 GM작물인 콩, 옥수수, 유채, 면화 및 벼에서 재배용으로 승인된 이벤트는 지속적으로 증가하여 2022년 10월 현재 274건에 달한다. 작물별 재배 승인 건 수는 GM옥수수가 135건으로 가장 많고, 면화 57건, 콩 40건, 유채 35건 및 벼 7건 순이다(Fig. 1, Table 1). 지난 삼십여 년 간 GM작물의 이벤트는 양적으로 증가하였을 뿐 아니라 질적 측면에서도 증가하였다. GM작물의 초기 도입형질은 잡초, 병해충과 같은 생물적 스트레스에 대응하여 작물보호와 재배 편의성을 높이거나, 유해 성분을 감소시켜 농업생산성을 높이는 것을 목적으로 개발되었다. 이 중 제초제 내성 형질은 가장 지배적인 도입형질로서 전세계 GM작물 재배면적의 45%를 차지하였으며, 재배된 주요 GM콩의 100%, GM옥수수와 유채는 90% 이상, GM면화의 73%에 제초제 내성 형질이 도입되었다 (Fig. 2A). 현재까지 주요 GM작물(콩, 옥수수, 유채, 면화 및 벼)에서 재배 승인된 274건의 이벤트 중 제초제 내성 형질이 포함된 이벤트는 총 213건으로, 대부분이 비선택성 제초제인 glyphosate 또는 glufosinate 내성 유전자를 포함하고 있으며, 그 외에 옥신계제초제인 dicamba, 2,4-D, PSII 저해제인 bromoxynil, ALS 저해제인 sulfonylurea계와 imazamox, HPPD 저해제인 isoxaflutole과 mesotrione 제초제 내성 유전자도 도입되었다(Fig. 2B). 비교적 최근인 2010년대 이후로는 기후변화에 대응하여 환경스트레스 내성을 높이거나 작물의 기능성 성분을 강화하는 방향으로 GM작물이 개발·승인되고 있다(ISAAA, 2018). 세부적인 도입형질의 종류는 작물 특성에 따라 다소 차이가 있는데 유지작물인 콩과 유채의 경우 지방산 조성을 조절한 이벤트가 추가되었으며, 타식율이 높은 옥수수와 유채는 웅성불임 유도 및 임성 회복 유전자를 이용한 수분 조절 형질이 빈번히 도입되었다(ISAAA, 2022).
|
Table 1. Traits and introduced genes of the major GM crops approved for cultivation up to 2022. 
|
|
zYear of the 1st event approved for cultivation in the trait category. |
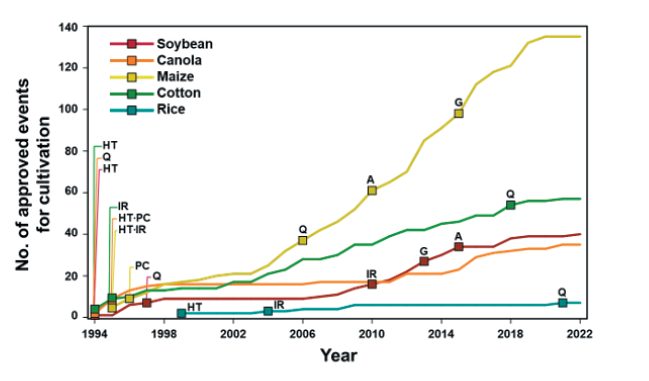
Fig. 1.Number of approvals for cultivation of major GM crops until 2022. The squares indicate the year when the novel trait was approved in each GM crops. HT, herbicide tolerance; IR, insect resistance; Q, modified product quality; PC, pollination system control; A, abiotic stress tolerance; G, altered growth/yield.
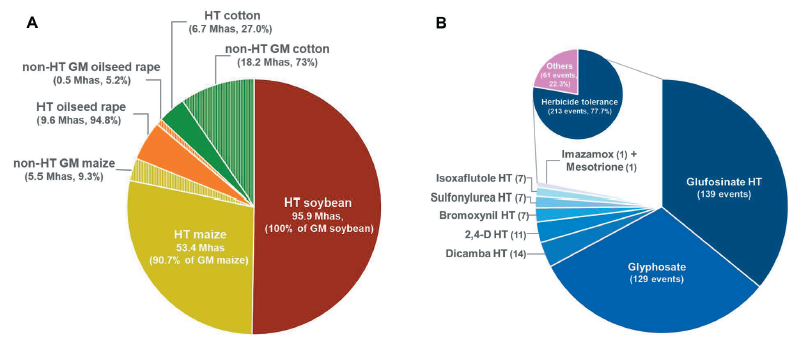
국내에서는 현재까지 GM작물의 상업적 재배가 승인되지 않았으며 사료용, 식품용 또는 산업용으로 수입 승인된 GM작물 이벤트는 7종의 작물에서 182건에 달한다(KBCH, 2022). 이 중 옥수수가 93건을 차지하였고, 면화 33건, 콩 29건, 유채 17건, 알팔파 5건, 감자 4건, 사탕무 1건이 뒤를 이었다. 수입 승인된 이벤트 중 단일 형질이 도입된 이벤트는 77건, 복합형질 이벤트는 105건이었는데, 이 중 85.2%는 제초제 내성 형질을 포함하고 있다. 이에 더하여 GM콩은 해충 저항성 및 영양성분 개선, GM유채는 임성 조절, 면화는 해충 저항성 관련 도입유전자를 지니고 있다. 국내 GM작물 수입량의 대부분(89.1%, 2021년)을 차지하는 옥수수는 제초제 내성, 해충 저항성, 영양성분 개선 형질 뿐만 아니라 최근에는 가뭄내성과 이삭중 증가 관련 형질을 포함하는 이벤트가 추가되었고, 다양한 교배조합을 통해 복합형질을 지닌 후대교배종이 다수 포함되어 있다(KBCH, 2021; 2022, Table 2).
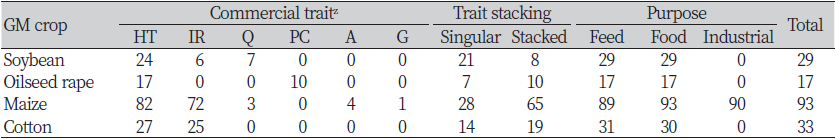
국내에서 GM 작물의 시험재배 등 환경방출을 위한 연구개발 승인은 벼, 콩, 고추, 잔디와 과채류의 형질전환 이벤트를 대상으로 2022년 9월 기준으로 총 4,151건에 달하며, 제초제 내성, 병해충 저항성, 기능성 물질 생산, 환경 스트레스 내성 및 고생산성 등의 형질에 대한 연구가 진행되고 있다. 특히, 주요 작물인 벼와 콩을 대상으로는 내냉성, 내서성, 내염성 및 내건성 등 다양한 환경 스트레스 내성 형질의 연구가 활발하다(KBCH, 2022)
주요 GM작물의 교잡특성 및 국내 근연종 분포
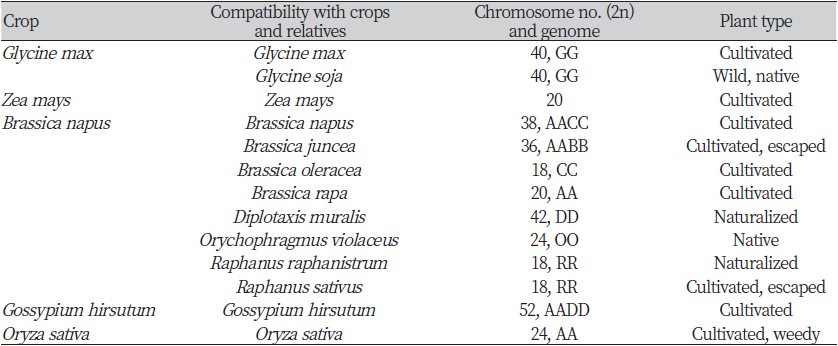
유전자 이입(introgression)은 한 유전자풀(gene pool)에서 다른 유전자풀로 유전자가 안정적으로 통합되는 것을 의미한다(Anderson, 1949). GM작물의 도입유전자가 생태계로 이입되려면 비변형(non-GM) 재배종과의 종내 교잡 또는 근연종과의 종간 교잡을 통해 임성을 지닌 후대를 생산해야 한다. 이를 위해서는 생식적 장벽(reproductive barrier)을 극복해야 하는데 교배 대상 사이의 화합성(compatibility), 개화 시기의 중첩, 수분매개체 등이 핵심 요인이다(Rieger et al., 1999; Umurzokov et al., 2021). 국내에서 주요 GM작물과 자연교잡이 발생할 가능성이 있는 국내 분포 근연종의 정보를 정리하여 Table 3에 제시하였다.
콩(Soybean)
재배콩(Glycine max; GG genome; 2n = 4× = 40)은 자식성 작물로 타식율이 1% 미만으로 알려져 있다(Carlson and Lersten, 2004; Caviness, 1966; Poehlman, 1987). 재배콩의 주두는 개화 약 24시간 전부터 개화 후 48시간 동안 화분을 수용할 수 있으며, 화분의 대부분은 자화수분한다(OECD, 2006). 일부 연구에서는 꿀벌(Apis mellifera)과 꽃노랑총채벌레(western thrips, Frankliniella occidentalis)이 재배콩의 타가수분에 기여하는 것으로 밝혀졌다(Chiari et al., 2005; Lee et al., 2015; Nakayama and Yamaguchi, 2002). 재배콩과 일년생 야생콩인 돌콩(Glycine soja; GG genome; 2n = 4× = 40)과 일년생 반야생콩인 G. gracilis (GG genome; 2n = 4× = 40)는 같은 Soja 아속에 속하며 염색체 수가 동일하여 종간 교잡이 가능하다(Yook et al., 2021; Karasawa, 1952). 재배콩의 직접적인 조상으로 여겨지는 돌콩은 중국 동부, 한국, 일본 및 러시아 극동 지역에 분포하고 있으며(Skvortzow, 1927; Lu, 2004; Zhuang, 1999), 재배콩과 돌콩의 중간 형태인 G. gracilis는 중국 동북부에서 관찰된다(Skvortzow, 1927).
국내에는 다양한 품종의 재배콩이 경작되며, 야생종으로는 돌콩이 전국적으로 분포한다(NIBR, 2022). 일반적으로 재배콩과 야생콩은 자가화합성이 우세하므로 자연교잡의 빈도가 매우 낮은 것으로 알려져 있으나 수정 후 장벽(postzygotic barrier)에 의한 생식적 격리가 없기 때문에 종간 교잡이 일어날 가능성이 충분하며, 한 번 유전자 이동이 발생한 개체가 생태계 내에서 살아남을 경우 자식에 의해 계속해서 도입유전자를 포함한 개체를 생산하며 장기간 지속될 우려가 크다(OECD, 2000)
옥수수(Maize)
옥수수(Zea mays; 2n = 2× = 20)는 타식성 식물로 주로 풍매로 번식한다, 화분의 양이 수술 하나에서 1800만립 이상으로 매우 많고 암이삭 하나에 낟알이 많이 달리는 특성이 있어 서로 다른 재배 품종들 사이의 교잡이 매우 쉽다(Kiesselbach, 1980; OECD, 2003). Zea 속에 속하는 야생종을 테오신테(teosinte)라 통칭하는데, 옥수수는 아종인 일년생 테오신테(Zea mays subsp. mexicana; 2n = 2× = 20)와 쉽게 종간 교잡하여 번식력 있는 후대 종자를 생산한다. 또한 매우 드물지만 Tripsacum 속(T. dactyloides; 2n = 72, T. floridanum; 2n = 36, T. lanceolatum; 2n = 72, T. pilosum; 2n = 72)과 속간 교잡이 가능하나 그 후대 종자는 높은 비율로 불임이며 유전적으로 불안정한 것으로 알려져 있다(Mangelsdorf, 1974).
우리나라 전역에서 다양한 품종의 옥수수를 재배하므로 GM옥수수가 재배된다면 주변의 비변형 옥수수 품종과 자연 교잡이 발생할 가능성이 높다. 다만 옥수수는 오랜 순화 과정을 통해 야생에서 생존할 수 있는 능력을 잃었고, 옥수수 종자는 암이삭에 달린 채 여러 겹의 포엽에 싸여 있어 탈립성이 극히 낮아 인간의 도움 없이 종자를 확산시키지 못하기 때문에 교잡후대종이 잡초화되어 생태계에 지속하기 어렵다(OECD, 2003). 한편 GM옥수수와 교잡 가능성이 있는 야생 근연종은 국내에 분포하지 않는다(NIBR, 2022).
유채(Oilseed rape)
유채(Brassica napus)는 38개의 염색체를 가지는 복2배체 식물로 AACC의 유전자형을 지니고 있으며, B. oleracea (AA genome; 2n = 2× = 18)와 B. rapa (CC genome; 2n = 2× = 20) 간 종간 교잡에 의해 유럽 대서양 또는 지중해 연안에서 기원한 것으로 추정된다(Liu, 1985). 유채는 원칙적으로는 자가수분 식물로 분류되나 타식율이 약 20-40%로 상당히 높은 것으로 알려져 있다(Becker et al., 1992). 화분의 양이 헥타르 당 1조립 정도로 매우 많아 풍매로 번식하며, 바람 외에도 꿀벌 등 곤충에 의한 수분 매개도 이루어진다(OECD, 2012; Zhang et al., 2018a). 유채가 속한 배추과(Brassicaceae)는 338속 3709종 이상의 식물종이 속해 있으며 종속간 교잡이 비교적 빈번한 것으로 알려져 있다. 인공수분 또는 실제 포장에서의 자연교잡에 의해 유채와의 종속간 교잡이 보고된 바 있는 배추과 식물은 19종에 달하였는데, Brassica 속의 8종(B. carinata, B. fruticulosa, B. juncea, B. maurorum, B. nigra, B. oleracea, B. rapa, B. tournefortii)과 종간 교잡이 보고되었고. Diplotaxis 속 3종(D. catholica, D. erucoides, D. muralis), Eruca sativa, Erucastrum gallicum, Hirschfeldia incana, Raphanus 속 2종(R. raphanistrum, R. sativus), Sinapis 속 2종(S. alba, S. arvensis), Orychophragmus violaceus 등 11종과 속간교잡이 보고되었다(OECD, 2012; Kim et al., 2018).
국가표준식물목록에 따르면 국내에는 38속 101종의 배추과 식물이 분포한다(Korea National Arboretum, 2022). 이 중 GM유채와 교잡 가능성이 있는 식물종은 8종이 있다(Table 3). 유채는 우리나라 원산은 아니지만 경관용, 채소 및 유지용 작물로 널리 재배되고 있으며, 갓(B. juncea), 배추(B. rapa), 꽃양배추(B. oleracea) 및 무(R. sativus)는 채소 작물로 전국적으로 재배된다. 배추, 꽃양배추와 무는 재배 특성상 개화 이전에 수확이 완료되어 GM유채와의 교잡 가능성이 희박하나 유채의 경우 지역 축제의 소재로 널리 활용되고 있으며 갓은 일부 야생화되어 GM유채와 교잡 가능성이 매우 높다. 귀화식물인 모래냉이(D. muralis)는 제주도에 서양무아재비(R. raphanistrum)는 안산과 인천 등지에 제한적으로 서식하며(NIBR, 2022), 자생종인 소래풀(O. violaceus)은 인공교배를 통해 정상 임성을 가지는 교잡 후대를 생산한 것이 보고되었다(Kim et al, 2018).
면화(Cotton)
목화속(Gossypium spp.)에는 약 50여 종이 속해 있는데, 45종은 2배체이고 5종은 4배체이다. 4개의 재배종(G. hirsutum, G. barbadense, G. aboreum, G. herbaceum)이 독립적으로 순화되었으며, 이 중 중미에서 순화된 이질4배체 재배종인 G. hirsutum (AADD genome; 2n = 4× = 52)은 전세계 면화 생산의 95%를 차지하며, 이를 모품종으로 GM면화가 개발되었다. 면화는 대부분 자가 수분하지만, 적절한 수분매개충이 있는 경우 타가수분도 가능하며, 타식율은 5-40%으로 변이폭이 크다. 면화의 화분은 크고 무거우며 약간 끈적하기 때문에 바람을 매개로 한 수분은 거의 일어나지 않는다(Kwon & Kim, 2001; OECD, 2008). 면화(G. hirsutum)는 다른 4배체 종(G. mustelinum, G. darwinii, G. barbadense, G. tomentosum)과 쉽게 종간 교잡하여 번식력 있는 후대 종자를 생산한다(OECD, 2008).
면화는 우리나라 전역에서 재배 가능하므로 GM목화와 자연 교잡될 가능성이 있다. 그러나 국내 목화 재배는 거의 전무한 실정이며, 설사 교잡이 일어난다 하여도 종자가 목화솜으로 둘러싸여 있어 탈립성이 극히 낮으므로 교잡후대종이 생태계에 침투하기 어렵다. 또한 GM면화와 교잡 가능성이 있는 야생 근연종은 국내에 분포하지 않는다(NIBR, 2022).
벼(Rice)
벼는 우리나라에서 가장 중요한 작물로서 재배면적과 생산량이 많아 자급이 되고 있다. Oryza 속은 2종의 재배벼(O. sativa와 O. glaberrima; AA genome; 2n = 2× = 24)와 20종의 야생벼를 포함 총 22종으로 구성되어 있다. 재배벼는 폐화수정하는 특성을 지니고 있어 기본적으로 자식성 작물이나 동종(재배종 또는 잡초성 벼) 간 자연교잡이 일부 가능하다. Oryza sativa와 O. glaberrima 간 자연교잡은 낮은 확률로 발생 가능하며, 교잡에 의해 생성된 F1의 화분은 불임률이 높다. Oryza sativa와 그 야생 조상으로 알려진 O. rufipogon (AA genome; 2n = 2× = 24) 간 교잡은 자연 상태에서 발생 가능하며, 교잡에 의해 생산되는 F1이 정상적인 임성을 가진다(OECD, 1999).
우리나라에는 자포니카 타입의 O. sativa만 재배되고 있으며 야생벼는 분포하지 않는 것으로 알려져 있다. 다만 재배벼와 야생벼의 중간 특성을 가지는 잡초성 벼인 앵미(red rice, O. sativa)가 존재하므로 GM벼로부터 잡초성 벼로 도입유전자가 이동할 우려가 있다(Kim et al., 2004).
국내 생태계에서 주요 GM작물의 유전자이동성 연구
종자 매개 유전자이동성 연구
생태계 내에서 GM작물의 도입유전자는 화분 또는 종자 형태로 이동 가능하다(Liu et al., 2013). GM옥수수와 면화는 국내에 자연교잡 가능한 야생종 또는 잡초종이 분포하지 않고 탈립성이 극히 낮은 특성으로 인해 화분을 통한 유전자이동에 의해 후대를 생산하여 생태계로 확산될 가능성이 낮을 것으로 예상되며, 실제로 국내에서 수행된 화분에 의한 유전자이동성 연구 사례를 찾아볼 수 없었다. 다만 사료용 및 식품용으로 수입된 종자의 비의도적 환경방출을 모니터링한 연구가 보고되었다. GM옥수수는 주로 사료용으로 수입되어 항만에서 사료가공 공장까지 운송되는 경로 및 축사 주변에서 비의도적으로 환경방출된 개체와 종자가 주로 발견되었다(Kim et al., 2006; Park et al., 2010; Han et al., 2014). 식물체의 경우 대체로 생육이 불량하여 번식 단계에 도달하지 못하였기 때문에 생태계 위해성이 낮은 것으로 판단되었으나 일부 축사 주변에는 비변형 옥수수 재배지가 있어 유전자이동이 발생할 우려가 있다(Park et al., 2010). GM면화 종자 역시 수입 후 운송 경로를 따라 주로 도로변이나 축산 농가 주변에서 발견되었다(Lim et al., 2021).
화분 매개 유전자이동성 연구
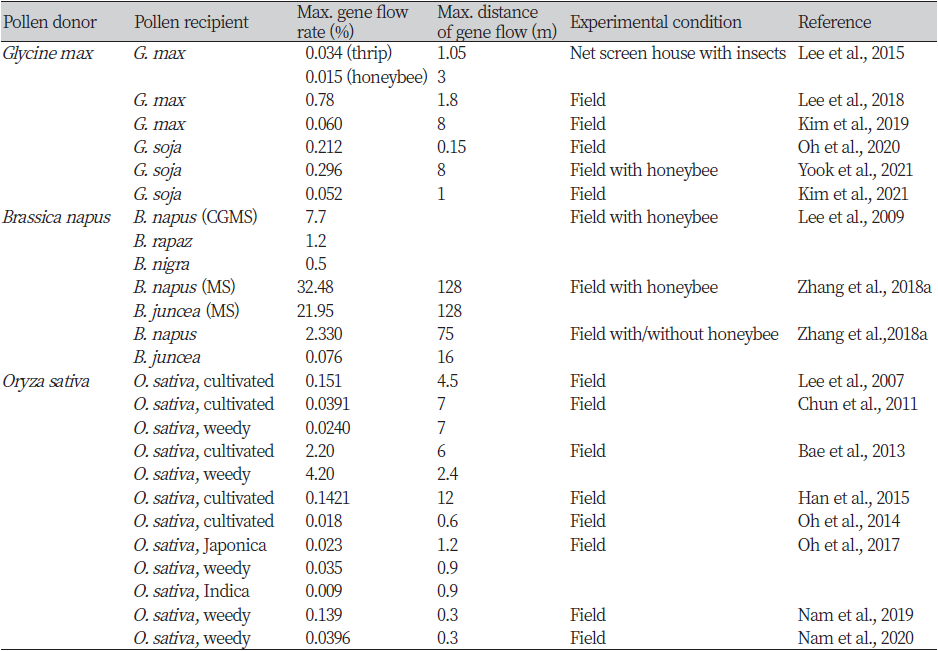
국내에 자연교잡 가능한 재배종, 근연 야생종 또는 잡초종이 분포하는 콩, 유채, 벼의 경우 이들의 GM작물에서 화분에 의한 유전자이동성 연구가 다수 보고되었다(Table 4). GM콩의 화분 매개 유전자이동성 연구는 재배종 또는 일년생 야생종인 돌콩을 대상으로 수행되어 자연교잡이 가능함이 실험적으로 증명되었으며, 교잡율에 다소 차이는 있으나 전반적으로 거리가 멀어질수록 유전자이동이 감소하는 추세를 나타냈다. 국내 야외포장 실험에서 GM콩-재배콩 간 자연교잡율은 0.15 m 거리에서 0.78%로 가장 높았으며(Lee et al., 2018), 최장 8 m (0.034%)까지 유전자이동이 발생하였다(Kim et al., 2019). Lee 등(2015)은 망실에서 GM콩-재배콩 간 유전자이동에 미치는 화분매개충의 영향을 연구하여 총채벌레는 근거리(0.15~1.05 m)에서의 GM콩 화분을 매개했을 가능성을 제시하였고, 꿀벌은 비행 특성에 따라 비교적 먼 거리(3 m)까지 화분을 이동시켰음을 보고하였다. 국내 야외포장 실험에서 GM콩-돌콩 간 자연교잡은 혼합재배(0 m) 조건에서 0.296%로 최대였고 최장 8 m 거리에서 0.025%의 교잡율을 나타냈다(Yook et al., 2021). 또한 GM콩-야생콩 간 교잡후대는 양친의 유전적 배경을 나누어 가지게 되므로 GM콩에 비해 종자생산성, 탈립성 및 휴면성 등이 강화되어 생태계 적응성이 확연히 높아지는 것이 확인되었다(Park et al., 2016; 2018; Yook et al., 2021).
유채는 배추과 내에서 종속간 교잡이 비교적 빈번한 것으로 알려져 있으며, 국내에서도 다양한 배추과 작물 사이의 종속간 교잡 연구가 이루어지고 있다(Kim et al., 2012; Lee et al., 2015; Seo et al, 2017; Kim et al., 2018). 국내 농업환경 조건에서 수행한 야외포장 실험에서 GM유채와 근연종 간 최대 자연교잡율은 비변형 유채, 갓, 배추 및 무에서 각각 2.33%, 0.076%, 1.2% 및 0%로 보고되었다(Lee et al., 2009b; Zhang et al., 2018b). Zhang 등(2018a)은 각각 방임수분(open pollination)과 풍매수분(wind pollination) 조건에서 거리에 따른 GM유채-근연종 간 유전자이동을 평가하여 바람에 의한 풍매수분과 꿀벌 등에 의한 충매수분의 비율을 정량적으로 규명하였다. GM유채로부터 비변형 유채와 갓으로의 근거리 화분 매개에는 꿀벌에 의한 충매의 역할이 더 컸으며, 원거리(유채 64 m 이상, 갓 32 m 이상) 화분 이동은 풍매의 역할이 다소 컸다. Lim 등(2015)은 생태계 내 지속성과 밀접한 특성인 종자의 월하/월동성 및 휴면특성을 평가하여 GM유채-갓 교잡종이 양친에 비해 높은 휴면성을 지녀 잡초화가능성이 높음을 보고하였다.
국내에서 GM벼의 화분 매개 유전자이동성 연구는 재배벼 또는 잡초성 벼(red rice, 앵미)를 대상으로 수행되었다. 실험 결과를 종합하면 GM벼와 자포니카 재배벼 사이의 자연교잡율은 최대 2.20%, 자연교잡이 발생한 최장 거리는 12 m에 달하였으며, GM벼-인디카 재배벼 간 최대 교잡율은 0.009%, 최장 거리는 0.9 m였고, GM벼-잡초성 벼에서는 각각 4.20%, 7 m였다(Table 4). 제초제 선발만으로 유전자 이동을 평가한 Bae 등(2013)의 연구에서 가장 높은 교잡율이 보고되었으며, 제초제 선발 및 분자마커 검정을 통해 유전자 이동을 확정한 다른 연구에서는 GM벼-재배벼 간 최대 0.151%, GM벼-잡초성 벼 간 최대 0.139%의 유전자 이동율이 보고되었다(Lee et al., 2007; Nam et al., 2018). 한편 지배적인 바람의 방향에 대칭으로 놓인 실험구에서 가장 높은 유전자이동이 발생하고, 개화기에 집중적인 강우가 겹칠 경우 자연교잡율과 유전자이동 거리가 모두 급감하는 것으로 보아 GM벼의 유전자이동성은 풍향과 강우 등의 기상 조건에도 크게 영향을 받음을 확인하였다(Bae et al., 2013; Han et al., 2015; Oh et al., 2017).
결론
본 리뷰는 다양한 연구 사례 및 정보를 수집·분석하여 국내 생태계에서 주요 GM작물의 자연교잡, 즉 화분을 매개로 하는 도입유전자의 이동 가능성을 제시하였다. GM작물의 화분 매개 유전자이동에는 다양한 요인이 관여하는데, 핵심요인은 교배 대상과의 화합성이다. 본 리뷰에 따르면 GM옥수수와 GM면화는 국내 생태계에서 자연교잡 가능한 근연 야생종 또는 잡초종이 없으므로 상대적으로 교잡가능성이 낮을 것으로 예상된다. 반면 GM콩, GM유채, GM벼는 전국에 야생종 또는 잡초종이 자생하며, GM작물과 개화기가 충분히 중첩되고 적합한 수분매개체가 존재하므로 국내 생태계에서 자연교잡에 의해 도입유전자가 확산될 가능성이 높다. GM작물과 근연종의 자연교잡연구 시에는 파종 시기를 조절하여 개화기를 맞추고, 실제 생태계보다 많은 수의 식물체와 수분 매개충을 투입하는 등 유전자 이동성을 극대화시키는 조건에서 실험이 이루어지므로 실제 생태계에서 GM작물과 근연종 간 자연교잡의 빈도는 훨씬 낮을 수 있다. 그럼에도 불구하고 GM콩, GM유채, GM벼는 자식하는 교잡특성에 의해 후속세대로 도입유전자가 지속적으로 유지 확산되어 생태계에 더 큰 영향을 줄 것으로 우려된다.
초기 도입형질인 제초제 내성, 영양성분 개선 등의 특성은 선택압력이 없는 경우에는 생태계에 위협이 되지 않았으나 도입형질에 따라 GM작물 이벤트의 생태계에 미치는 위해도 편차가 존재한다. 최근 개발되는 환경 스트레스 내성 및 광합성 효율 증대와 같은 도입형질에 대한 선택압력은 생태계에서 보편적으로 작용할 수 있으며, 이 경우 도입유전자 효과에 의해 GM작물이 높은 적응성을 가질 것이라 예측할 수 있다. 이후 자연교잡을 통해 해당 GM작물이 근연 야생종 또는 잡초종의 유전적 배경을 획득하면 생태계 내 적응성이 더욱 높아져 생태계를 교란시킬 우려가 크다. 따라서 국내 생태계에서 근연 야생종 또는 잡초종과의 자연교잡 가능성이 높은 GM콩, GM유채, GM벼에 대해서는 도입형질에 따라 차별화된 안전관리 전략의 수립이 필요할 것으로 판단된다.

